Анализ структуры тРНК и ДНК-белковых взаимодействий
Исполнитель: Михаил Сучков
1. Предсказание вторичной структуры тРНК (1G59)
Для последовательности тРНК 1G59 был выполнен поиск инвертированных повторов и расчет оптимальной вторичной структуры по алгоритму Зукера.
Результат предсказания (ViennaRNA):
Structure: (((((((..((((........))))(((((.......))))).....(((((.......))))))))))))..
Energy: -35.20 kcal/mol
Предсказанная структура полностью совпадает с классической моделью «клеверного листа».
2. Анализ контактов в комплексе 1DDN (EcoRI)
В ходе работы были исследованы взаимодействия между белком EcoRI (цепь D) и ДНК (цепи E, F). С помощью PyMOL проведен расчет полярных (до 3.5 Å) и неполярных (до 4.5 Å) контактов.
Распределение контактов по группам атомов
| Группа атомов ДНК | Полярные (< 3.5 Å) | Неполярные (< 4.5 Å) | Всего |
|---|---|---|---|
| Дезоксирибоза (сахара) | 4 | 11 | 15 |
| Фотифатные группы | 4 | 12 | 16 |
| Основания (Большая бороздка) | 1 | 6 | 7 |
| Основания (Малая бороздка) | 0 | 0 | 0 |
3. Визуализация ключевых аминокислотных остатков
3.1 Остаток с наибольшим числом контактов (Lys113)
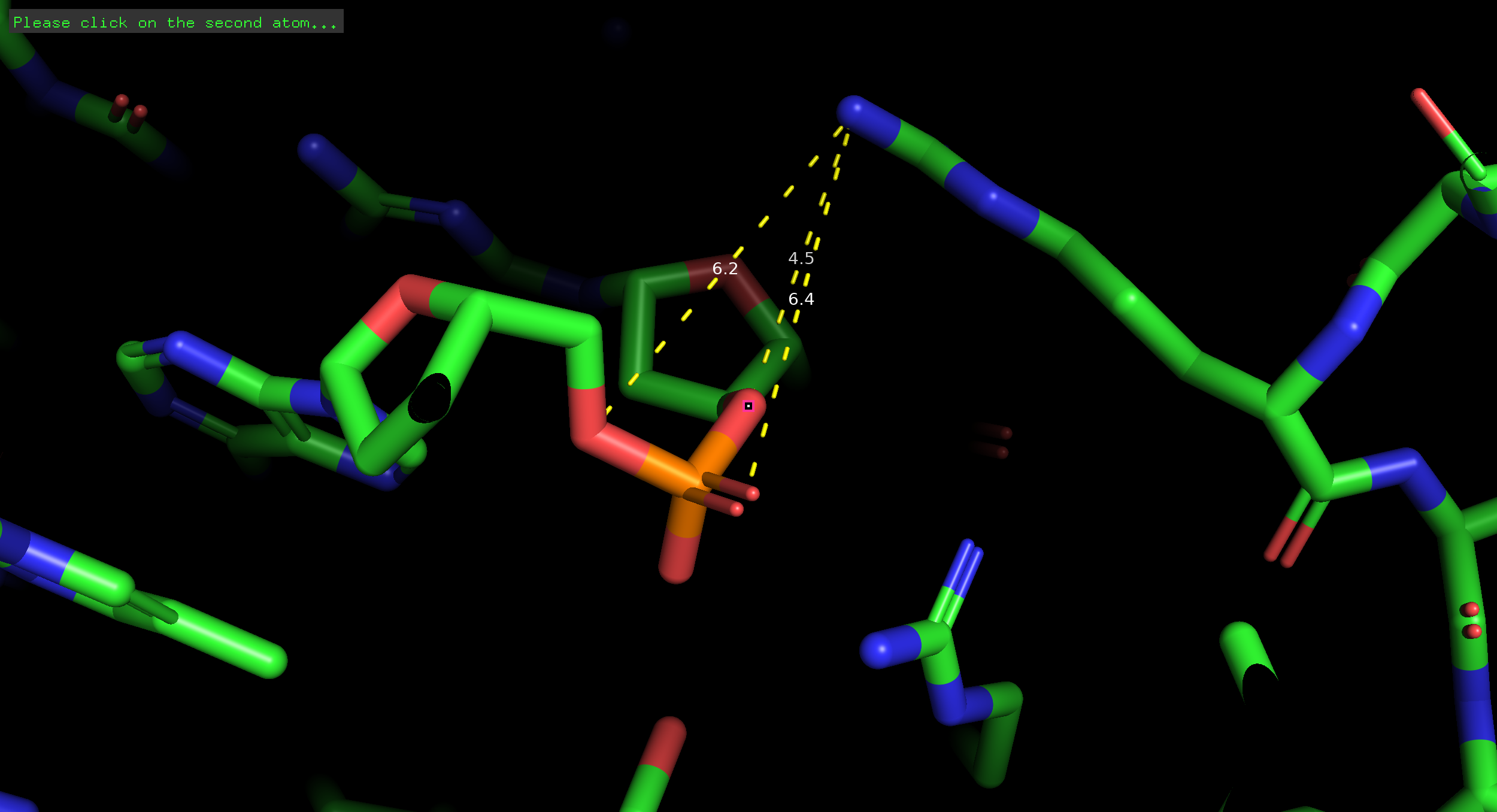
Рисунок 1. Взаимодействие Lys113 с фосфатным остовом ДНК.
Обоснование: Остаток Lys113 образует максимальное количество контактов с остовом ДНК. Его положительно заряженная боковая группа взаимодействует с отрицательно заряженными кислородами фосфатных групп. Это обеспечивает неспецифическую стабилизацию комплекса («якорная» функция).
3.2 Остаток, важный для распознавания (Arg145)
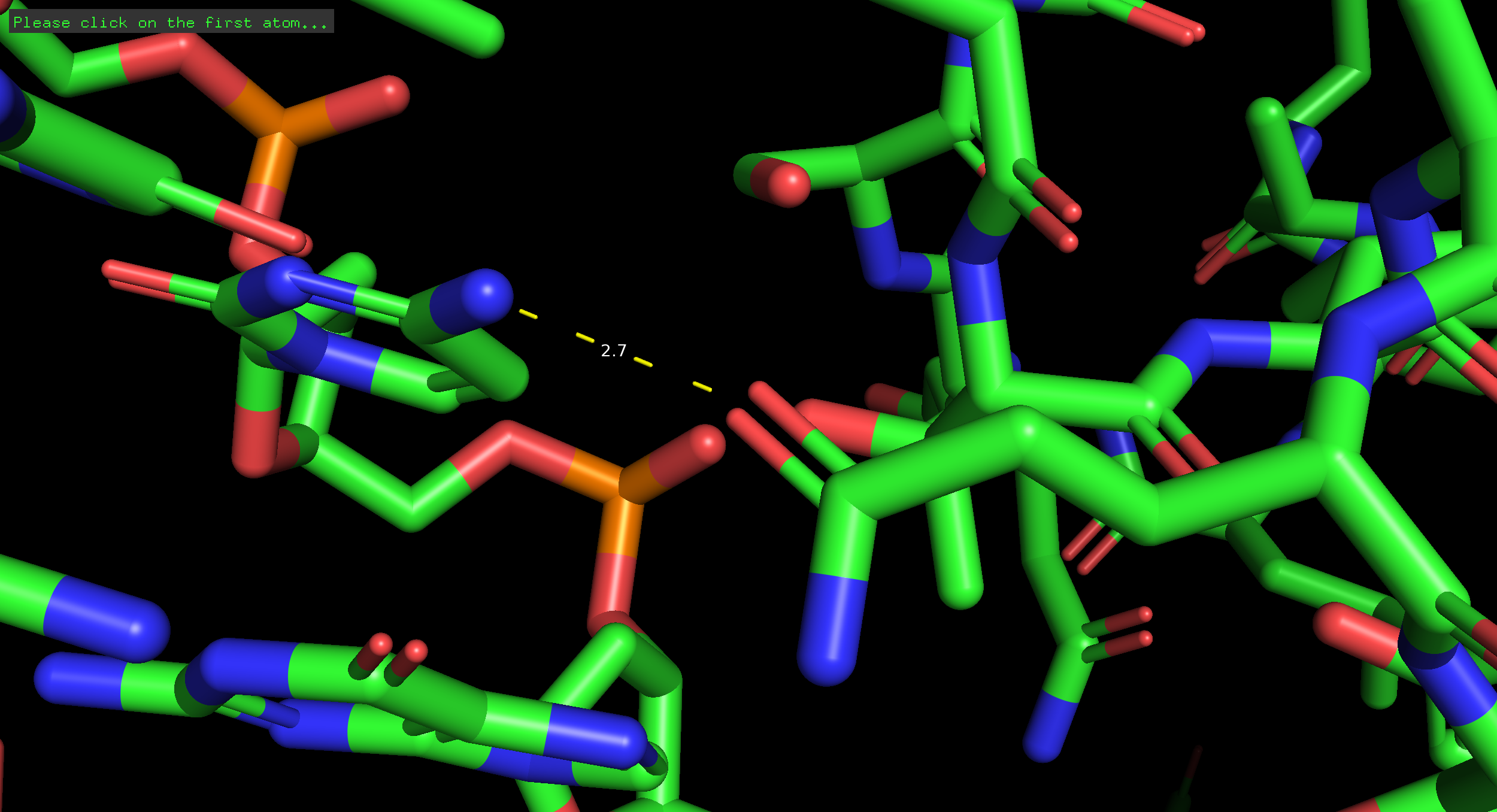
Рисунок 2. Специфические связи Arg145 в большой бороздке.
Обоснование: Остаток Arg145 является ключевым для специфического узнавания последовательности GAATTC. Он проникает в большую бороздку ДНК и образует водородные связи непосредственно с азотистыми основаниями, считывая генетическую информацию.
Выводы
Анализ структуры 1DDN показал, что белок EcoRI использует два типа стратегий связывания: электростатическое взаимодействие с остовом для удержания ДНК и специфическое взаимодействие в большой бороздке для узнавания целевого сайта рестрикции.