Главная страница > Четвертый семестр > Коды ферментов
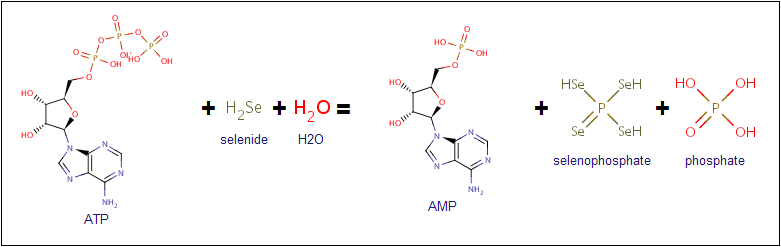
Информация о значении кода фермента EC 2.7.9.3 была получена на сайте IUPAC. Первая цифра кода (2) указывает на то, что фермент является трансферазой, то есть участвует в переносе какой-либо группы с одной молекулы на другую. Вторая (7) и третья (9) цифры кода фермента обозначают переносимую группу и тип акцептора переносимой группы соответственно. В данном случае имеет место перенос фосфата, в качестве акцепторов выступают две молекулы (то есть одновременно фосфорилируются два субстрата). Четвертая цифра обозначает сам фермент (катализируемую реакцию) — селенид, H2O-дикиназу (selenid, water-dikinase; селенофосфатсинтетаза). Селенофосфатсинтетаза осуществляет перенос двух фосфатных групп ATP на селенид и H2O с образованием AMP, неорганического фосфата и селенофосфата.
Рассмотрены последовательности селенофосфатсинтетаз (EC 2.7.9.3) из организмов Escherichia coli K-12, Methanococcus jannaschii и Homo sapiens. Последовательности ферментов были получены с помощью следующего запроса SRS:
([uniprot-ECNumber:2.7.9.3] & (([uniprot-ID:*_ecoli] | [uniprot-ID:*_metja]) | [uniprot-ID:*_human]))
Результаты поиска приведены здесь (выбран способ отображения SW_InterProMatches). Были найдены две последовательности селенофосфатсинтетаз человека (кодируются генами SPS1 и SPS2, UniProt ID SPS1_HUMAN и SPS2_HUMAN), по одной последовательности селенофосфатсинтетаз-2 E.coli K-12 и M.jannadchii (кодируются генами selD, UniProt ID SELD_ECOLI и SELD_METJA) и трансляты двух человеческих cDNA (UniProt ID Q8N9T3_HUMAN и Q8NAW0_HUMAN), которые, по-видимому, синтезированы на мРНК Sps2 (см. выравнивание последовательностей Sps2 человека и двух cDNA). Для дальнейшего рассмотрения отобраны первые четыре последовательности.
Все последовательности имеют одинаковую доменную структуру Pfam: содержат N-концевой домен AIRS (PF00586) и C-концевой домен AIRS_C (PF02769). По данным InterPro, в большинстве случаев оба домена входят в состав последовательностей белков одновременно; некотрые бактериальные фосфорибозилформилглицинамидинсинтазы (EC 6.3.5.3, например, из организмов Shewanella sp., Polynucleobacter sp. и Burkholderia cenocepacia) содержат один домен AIRS и два домена AIRS_C. Белки, имеющие данные домены, участвуют в различных ферментативных реакциях, общей чертой которых является гидролиз ATP. При этом может иметь место фосфорилирование таких субстратов, как Se2- и интермедиаты биосинтеза пуринов: 2-N-формил-1-N-(5-фосфо-D-рибозил)глицинамид, 2-(формамидо)-1-N-(5-фосфо-D-рибозил)ацетамидин и H2O — ATP-зависимая дегидратация (R-C(O)NH2 = R-CN + H2O). При этом домен AIRS предположительно содежит ATP-связывающий центр, функция домена AIRS_C неизвестна. Информация о молекулярном механизме реакций, осуществляемых белками, содержащими домены AIRS и AIRS_C, взята из БД EcoCyc.
Информация о доменной структуре селенофосфатсинтетаз приведена в табл. 2. В связи с тем, что рассматриваемые белки состоят из гомологичных доменов (имеющих одинаковое расположение), их также можно считать гомологичными.
Для оценки попарного сходства гомологичных доменов последовательности белков, найденных с помощью SRS, были сохранены в FASTA-формате. Затем с помощью команды seqret были получены последовательности доменов и проведено их множественное выравнивание с использованием программы muscle (Edgar, 2004). Алгоритм данной программы позволяет получать наиболее точные выравнивания (по сравнению с другими распространенными алгоритмами: T-Coffee и ClustalW), что существенно при выравнивании последовательностей из значительно удаленных друг от друга организмов. Построенные выравнивания приведены здесь (красным отмечены позиции, содержащие четыре одинаковых аминокислотных остатка, зеленым — три, желтым — два).
Измерение попарного сходства (similarity) между последовательностями гомологичных доменов было проведено на основе построенных выравниваний с помощью программы fprotdist (программа запущена с параметром -method s). Выдачи программы для доменов AIRS и AIRS_C приведены на рис. 1 и 2 соответственно (значения попарного сходства были переведены из долей в проценты). Работа с программами была осуществлена с помощью следующего скрипта Linux:
seqret seld_ecoli.fasta 1_1.fasta -sbegin1 7 -send1 158 -sreverse N
seqret seld_metja.fasta 1_2.fasta -sbegin1 11 -send1 161 -sreverse N
seqret sps1_human.fasta 1_3.fasta -sbegin1 32 -send1 188 -sreverse N
seqret sps2_human.fasta 1_4.fasta -sbegin1 52 -send1 239 -sreverse N
seqret seld_ecoli.fasta 2_1.fasta -sbegin1 169 -send1 345 -sreverse N
seqret seld_metja.fasta 2_2.fasta -sbegin1 170 -send1 341 -sreverse N
seqret sps1_human.fasta 2_3.fasta -sbegin1 192 -send1 368 -sreverse N
seqret sps2_human.fasta 2_4.fasta -sbegin1 243 -send1 418 -sreverse N
cat 1_1.fasta > domain_1.fasta
cat 1_2.fasta >> domain_1.fasta
cat 1_3.fasta >> domain_1.fasta
cat 1_4.fasta >> domain_1.fasta
cat 2_1.fasta > domain_2.fasta
cat 2_2.fasta >> domain_2.fasta
cat 2_3.fasta >> domain_2.fasta
cat 2_4.fasta >> domain_2.fasta
rm 1_1.fasta
rm 1_2.fasta
rm 1_3.fasta
rm 1_4.fasta
rm 2_1.fasta
rm 2_2.fasta
rm 2_3.fasta
rm 2_4.fasta
muscle -in domain_1.fasta -out domain_1_align.msf -msf
muscle -in domain_2.fasta -out domain_2_align.msf -msf
fprotdist domain_1_align.msf domain_1.fprotdist -method s -auto
fprotdist domain_2_align.msf domain_2.fprotdist -method s -auto
Попарное сходство между последовательностями доменов AIRS и AIRS_C белков Sps2 и Sps2 человека составляет 75% и 86% соответственно, тогда как попарное сходство между любыми другими последовательностями не превышает 41%. По результатам Guimarães et al. (1996), как Sps1, так и Sps2 катализируют одну и ту же реакцию, но Sps1 экспрессируется в клетках взрослого организма, а Sps2 — преимущественно на ранних стадиях эмбриогенеза. При этом Sps1 и Sps2 участвуют в различных метаболических путях: синтез селенофосфата из селенита de novo и синтез селенофосфата из селена (в нулевой степени окисления) соответственно, по результатам Tamura et al. (2004). Таким образом, у человека имеется два паралога селенофосфатсинтетазы, которые разошлись значительно позже, чем эукариоты и бактерии и эукариоты и археи.
Несмотря на низкое попарное сходство (до 25% между последовательностями доменов AIRS_C Sps2 человека и SelD E.coli K-12), рассматриваемые ферменты катализируют одну и ту же реакцию. Следовательно, низкое сходство последовательностей белков не означает, что эти белки должны выполнять разные функции. Для выполнения белком его функции большее значение имеет не аминокислотная последовательность, а пространственная структура: белки со значительно отличающимися последовательностями могут иметь сходные пространственные структуры. Кроме того, высокая консервативность характерна для аминокислотных остатков активных центров, непосредственно принимающих участие в катализе (вероятно, группы из нескольких абсолютно консервативных позиций на выравниваниях образуют активные центры ферментов).
Табл. 1. Доменная структура UniProt-белков Escherichia coli K-12, Methanococcus jannaschii и Homo sapiens, имеющих EC 2.7.9.3.
| № | UniProt ID | UniProt AC | Имя гена | Первый домен | Второй домен | ||
| Идентификатор Pfam | Положение в последовательности | Идентификатор Pfam | Положение в последовательности | ||||
| 1 | SELD_ECOLI | P16456 | selD | PF00586 | 7-158 | PF02769 | 169-345 |
| 2 | SELD_METJA | P60819 | selD | PF00586 | 11-161 | PF02769 | 170-341 |
| 3 | SPS1_HUMAN | P49903 | SEPHS1 (SPS1, SELD) | PF00586 | 32-188 | PF02769 | 192-368 |
| 4 | SPS2_HUMAN | Q99611 | SEPHS2 (SPS2) | PF00586 | 52-239 | PF02769 | 243-418 |
SPS1_HUMAN SPS2_HUMAN SELD_ECOLI SELD_METJA
SPS1_HUMAN 100.00% 74.52% 31.16% 35.97%
SPS2_HUMAN 74.52% 100.00% 32.21% 35.76%
SELD_ECOLI 31.16% 32.21% 100.00% 40.41%
SELD_METJA 35.97% 35.76% 40.41% 100.00%
|
Рис. 1. Таблица попарного сходства (table of similarity) доменов AIRS (PF00586) селенофосфатсинтетаз из организмов Escherichia coli K-12, Methanococcus jannaschii и Homo sapiens.
SELD_ECOLI SELD_METJA SPS1_HUMAN SPS2_HUMAN
SELD_ECOLI 100.00% 28.57% 27.22% 24.84%
SELD_METJA 28.57% 100.00% 36.97% 40.00%
SPS1_HUMAN 27.22% 36.97% 100.00% 86.36%
SPS2_HUMAN 24.84% 40.00% 86.36% 100.00%
|
Рис. 2. Таблица попарного сходства (table of similarity) доменов AIRS_C (PF02769) селенофосфатсинтетаз из организмов Escherichia coli K-12, Methanococcus jannaschii и Homo sapiens.
Рассмотрены данные БД BRENDA, относящиеся к селенофосфатсинтетазам (EC 2.7.9.3). Схема катализируемой селенофосфатазами реакции приведена на рис. 3 (механизм реакции был установлен на примере селенофосфатсинтетаз E.coli и Haemophilus influenzae). В БД BRENDA описана 71 последовательность ферментов с EC 2.7.9.3 (селенофосфатсинтетаз). Данный фермент присутствует в клетке в виде мономеров (поле Subunit, Kim and Stadtman, 1995). Посттрансляционные модификации селенофосфатсинтетаз не описаны (поле Posttranslational Modification). Болезни, связанные с селенофосфатсинтетазами, также неизвестны (поле Disease). Селенофосфатсинтетаза играет ключевую роль в обеспечении функционирования всех селеноцистеинсодержащих ферментов. Возможно, нарушения, связанные с его функционированием, приводят к гибели на ранних стадиях эмбриогенеза и, таким образом, не проявляться позднее в виде болезней.
Рис. 3. Схема реакции, катализируемой селенофосфатсинтетазами. По данным BRENDA.
Рассмотрены аннотации последовательностей селенофосфатсинтетаз (EC 2.7.9.3) из организмов Escherichia coli K-12, Methanococcus jannaschii и Homo sapiens (UniProt ID SPS1_HUMAN, SPS2_HUMAN, SELD_ECOLI и SELD_METJA). В БД GOA ни с одной из перечисленных записей UniProt, кроме SPS2_HUMAN, не ассоциирован ни один из терминов онтологии Component (внутриклеточная локализация, четвертичная структура), а ассоциированный с SPS2_HUMAN термин GO:0005575 (компонент клетки, cellular component) не имеет надежных подтверждений (код подтверждения ND). Таким образом, локализация селенофосфатсинтетаз в клетке практически не изучена. С записями SPS1_HUMAN и SPS2_HUMAN (в отличие от SELD_ECOLI и SELD_METJA) ассоциировано по одному термину онтологии Process (процесс): GO:0006464 (модифицирование белков, protein modification, код подтверждения TAS) и GO:0016260 (биосинтез селеноцистеина, selenocysteine biosynthetic process, код подтверждения NAS) соответственно. По результатам Low et al. (1995), Sps1 человека принимает участие в синтезе селеноцистеил-tRNASec, поставляя необходимый для этого интермедиат — селенофосфат (ссылка на статью приведена в аннотации GOA). Селеноцистеил-tRNASec участвует в биосинтезе селеноцистеинсодержащих белков (например, глутатионпероксидазы). Таким образом, Sps1 также принимает участие в биосинтезе селеноцистеина, однако запись SPS1_HUMAN не ассоциирована с термином биосинтез селеноцистеина. Аналогично Sps2, как и Sps1, поставляет селенофосфат для биосинтеза селеносодержащих белков, однако запись SPS2_HUMAN не ассоциирована с термином модифицирование белков. Оба термина имеют коды подтверждения, указывающие на то, что они были присвоены куратором (NAS и TAS). Возможно, различия аннотаций записей SPS1_HUMAN и SPS2_HUMAN связаны с тем, что аннотация осуществлялась независимо двумя разными кураторами.
С каждой из записей UniProt ассоциировано не менее шести терминов онтологии Function (молекулярный механизм и специфичность). Большинство из терминов имеют коды подтверждения IEA, то есть были присвоены на основе поиска сходных последовательностей с известной функцией. Экспериментальное подтверждения имеют только термины, указывающие на способность Sps1 связывать ATP и ADP (Low et al., 1995) и на способность Sps2 осуществлять рекцию ATP + селенид + H2O = AMP + селенофосфат + Pi (Guimarães et al., 1996). Набор терминов GOA онтологии Function для каждой из записей UniProt совпадает, за исключением термина GO:0000287 (связывание ионов Mg2+, magnesium ion binding), ассоциированного только с прокариотическими записями, и GO:0005525 (связывание GTP, GTP binding), ассоциированного только с SPS1_ECOLI. По данным HAMAP, ион Mg2+ является кофактором селенофосфатсинтетазы. Отсутствие ассоциаций эукариотических записей UniProt с термином связывание ионов Mg2+ обусловлено тем, что данный термин был ассоциирован с прокариотическими записями на основе информации БД HAMAP, содержащей аннотации только прокариотических семейств белков и семейств белков пластид. Другие термины онтологии Function, ассоциированные с рассматриваемыми записями UniProt, указывают на способность селенофосфатсинтетаз связывать ATP (GO:0005524), связывать селен (GO:0008430) и катализировать реакцию ATP + селенид + H2O = AMP + селенофосфат + Pi (GO:0004756), или являются родительскими по отношению к данным терминам.
Аннотации прокариотических записей UniProt (UniProt ID SELD_ECOLI и SELD_METJA) практически совпадают с аннотациями GOA. Это связано с тем, что они были импортированы из одного источника (HAMAP) в обе БД. Единственным отличием является то, что в аннотациях GOA приведены некоторые родительские термины для терминов HAMAP, а в UniProt — нет. При этом все родительские термины были импортированы из других БД, в которых приведена менее подробная информация, чем в HAMAP (для ассоциирования терминов с записями UniProt GOA использует большее количество источников информации, чем UniProt). Аннотации UniProt эукариотических записей (UniProt ID SPS1_HUMAN и SPS2_HUMAN) менее подробны, чем аннотации GOA. Например, для Sps2 человека не указана способность связывать ATP и участие в биосинтезе селеноцистеина. Таким образом, аннотирование записей в UniProt отстает от аннотирования записей в GOA (GOA является специализированной БД, содержащей только аннотации, БД UniProt в первую очередь предназначена для хранения аминокислотных последовательностей).
Проведено сравнение свойств селенофосфатсинтетаз (EC 2.7.9.3) из организмов E.coli K-12 и Homo sapiens. Для этого была осуществлена фильтрация всех записей BRENDA, относящихся к селенофосфатсинтетазам, по принадлежности к данным организмам. Для селенофосфатсинтетазы E.coli, в отличие от селенофосфатсинтетаз человека, известно оптимальное значение pH (7.2). Единственным известным ингибитором селенофосфатсинтетаз человека является 8-азидо-ATP (конкурентный механизм ингибирования, рис. 4). Данный ингибитор, содержащий 32P, был использован для мечения селенофосфатсинтетазы-1 в работе Low et al. (1995). Было показано, что данное вещество, как и другие искусственные аналоги ATP (α, β-метилен-ATP и γ-тио-ATP) и АМP являются ингибитором селенофосфатсинтетазы E.coli (Veres et al., 1994). Кроме того, селенофосфатсинтетазу E.coli ингибируют H2O2 и различные неорганические ионы (Li+ и Na+ в присутствии K+, Zn2+ — конкурентно по отношению к кофактору селенофосфатсинтетазы Mg2+, фосфат и селенофосфат — продукты катализируемой реакции, смещающие равновесие влево). Возможно, данные ингибиторы действуют и на селенофосфатдегидрогеназы человека, что не было проверено экспериментально (большая изученность селенофосфатсинтетазы E.coli по сравнению с селенофосфатсинтетазой человека, скорее всего, связана с возможностью выделения в больших количествах). Активаторы селенофосфатдегидрогеназ неизвестны.
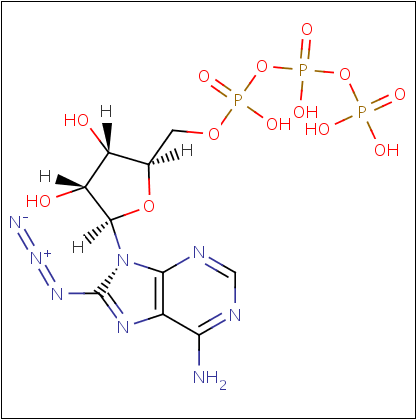
Рис. 4. 8-азидо-ATP, конкурентный ингибитор селенофосфатсинтетазы. По данным BRENDA.
© Куравский Михаил Львович, 2007