Сигналы в геноме. Примеры и поиск сигналов с известными последовательностями
Описание сигнала RSS
Краткое описание
Название сигнала: recombination signal sequence (сигнальнвая последовательность рекомбинации)
В чём состоит сингнал: данный сигнал необходим для V(D)J рекомбинации
Кому адресован: ферментативный комплекс RAG1/RAG2
Предназначение: белки RAG узнают сигнал и вносят одноцепочечный разрыв между сегментами V, D и J и их фланкирующими последовательностями RSS
Сила сигнала: белки RAG должны случайным образом связываться с одной из RSS между V-сегментами и с другой — между J-сегментами. При этом не должно быть такого, что в какой-то области комплекс не связался ни с одной RSS, иначе антитело не получится. Так что сигналы должны быть довольно сильными, а раз все RSS одинаковы по силе, выбор осуществуляется случайным образом.
Подробное описание
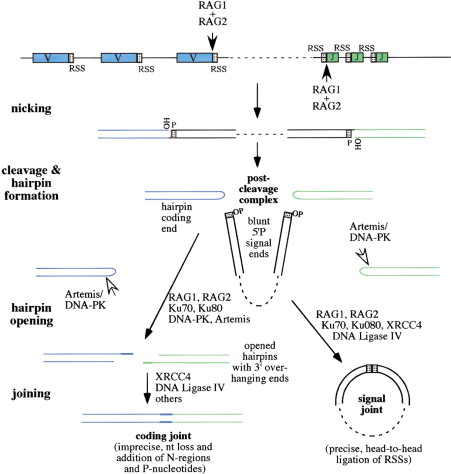
Для этого задания я выбрала сигнал RSS – recombination signal sequence – это фланкирующие последовательности сегментов V, D и J. V(D)J рекомбинация создает разнообразие в иммунной системе посредством лимфоид-специфичной сборки множества генных сегментов в функциональные гены иммуноглобулинов и рецепторов Т-клеток. V(D)J рекомбинация инициируется белками RAG1 и RAG2, которые вводят двухцепочечные разрывы ДНК между сегментами V (variable), D (diversity) и J (joining) и их фланкирующими сигнальными послеоваткльносятми RSS. Расположение кодирующих сегментов и RSS, а также схема V(D)J рекомбинации представлена на Рис 1. RSS состоят из 3 частей:
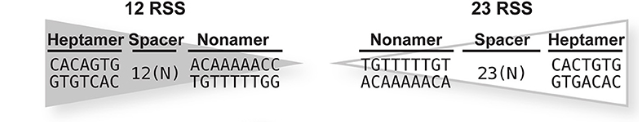
- консервативный гептамер
- спейсер из 12 или 23 пар оснований
- консервативный нонамер
RSS делят на 12-RSS и 23-RSS по длине спейсера. Сплайсинг может происходить только между одним сегментом, фланкированным 12-RSS, и другим сегментом, фланкированным 23-RSS, как гласит правило 12/23. Для оптимальной активности RAG необходимы взаимодействия как с консервативным гептамером, так и с нонамером. 12-RSS и 23-RSS связываются с одним и тем же RAG. Было показано, что белок группы с высокой подвижностью HMGB1 стимулирует ДНК-связывающую активность RAG, предположительно индуцируя изгиб RSS. RSS распознается обеими субъединицами RAG1 в димере RAG, при этом начальная часть в основном распознается инсерционным доменом (ID) и РНКаза H-подобным доменом (RNH) одной субъединицы, а более дистальная часть распознается RNH, ДНК-связывающим доменом и С-концевым доменом (CTD) симметричной субъединицы. Множенственные взаимодействия наблюдаются в первых трёх позициях гептамера, чем объясняется идеальная консервативность последовательности этих нуклеотидов в разных геномах (консенсусная последовательность RSS представлена на Рис 2). При этом основание первого нуклеотида выворачивается из дуплекса. Связывание одного RSS индуцирует конформационные изменения в обоих мономерах RAG1, и это может способствовать образованию парных комплексов 12-RSS и 23-RSS. Когда 12-RSS связывается как гептамером, так и нонамером с в HMGB1-изогнутой конформации, димер NBD наклоняется в сторону более короткого 12-RSS, оставляя тот же самый димер NBD со способностью связывания только с 23-RSS, также в HMGB1-изогнутой конформации. И наоборот, когда 23-RSS связан, димер NBD отклоняется в сторону от более длинного 23-RSS, оставляя тот же самый димер NBD со способностью взаимодействовать только с 12-RSS. Также каталитический остаток в активном центре находится на спирали, которая взаимодействует с RSS. Таким образом, связывание RSS может также индуцировать каталитические конформации в RAG1.
После внесения разрыва белками RAG образовавшаяся 3’-ОН группа атакует фосфодиэфирную связь в противоположной цепи. На концах кодирующих сегментов образуются ДНК-шпильки, которые узнаются белками Ku70/Ku80, а Artemis разрезает шпильки (несиквенсспецифично). Далее концы кодирующих сегментов соединяются по тому же механизму, что и при негомологичном соединении концов при репарации ДНК. Так остаются только определённые V, D и J сегменты. Перед этим терминальная дезоксинуклеотидил трансфераза (TdT) достраивает несколько нуклеотидов (до 10-20) случайным образом, что дает дополнительную вариабельность. Вырезанный участок, содержащий сигнальные последовательности RSS, замыкается с образованием кольцевидной структуры, известной как рекомбинационное вырезанное кольцо.
Литература
- Mark S. Schlissel "Does Artemis End the Hunt for the Hairpin-Opening Activity in V(D)J Recombination?" doi:10.1016/S0092-8674(02)00694-3
- Heng Ru, Melissa G. Chambers, Tian-Min Fu, Alexander B. Tong, Maofu Liao, and Hao Wu "Molecular Mechanism of V(D)J Recombination from Synaptic RAG1-RAG2 Complex Structures" doi: 10.1016/j.cell.2015.10.055
- Uthayashanker R. Ezekiel,Tianhe Sun,Grazyna Bozek &Ursula Storb "The Composition of Coding Joints Formed in V(D)J Recombination Is Strongly Affected by the Nucleotide Sequence of the Coding Ends and Their Relationship to the Recombination Signal Sequences" https://doi.org/10.1128/MCB.17.7.4191
- Craig H. Bassing, Frederick W. Alt, Maureen M. Hughes, Margaux D'Auteuil, Tara D. Wehrly, Barbara B. Woodman, Frank Gärtner, J. Michael White, Laurie Davidson and Barry P. Sleckman "Recombination signal sequences restrict chromosomal V(D)J recombination beyond the 12/23 rule" Nature 405, 583–586 (2000) https://doi.org/10.1038/35014635
- Dale A. Ramsden, Tanya T. Paull & Martin Gellert "Cell-free V(D)J recombination" Nature 388, 488–491 (1997) https://doi.org/10.1038/41351
- Alastair L. Smith, James N. F. Scott, Joan Boyes "The ESC: The Dangerous By-Product of V(D)J Recombination" https://doi.org/10.3389/fimmu.2019.01572
Построение PWM для последовательности Козак человека
Для выполнения этого задания я использовала скрипт, позаимствованный у наших уважаемых старшекурсников.
Он строит PWM для последовательностей следующего вида:
7 нуклеотидов до ATG + ATG + 3 нуклеотида после ATG
В результате работы скрипта получаются 3 выборки: 100 последовательностей для построения PWM, 200 последовательностей для тестирования, где ожидается сигнал и 200 последовательностей для отрицательного контроля со случайными ATG.
Полученная PWM с псевдоотсчётами представлена в Таблице 1.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | -0,44 | -0,61 | -0,67 | -0,17 | 0,28 | -0,25 | -0,44 | 1,21 | -5,00 | -5,00 | -0,17 | 0,02 | -0,89 |
| T | -0,25 | -0,34 | -0,25 | -0,49 | -1,29 | -0,81 | -1,74 | -5,00 | 1,21 | -5,00 | -0,98 | -0,49 | -0,17 |
| G | 0,44 | 0,64 | 0,41 | 0,20 | 0,67 | 0,38 | 0,38 | -4,64 | -4,64 | 1,58 | 0,80 | -0,18 | -0,64 |
| C | 0,24 | 0,16 | 0,41 | 0,44 | -0,45 | 0,50 | 0,80 | -4,64 | -4,64 | -4,64 | -0,13 | 0,53 | 0,16 |
Гистограммы весов:
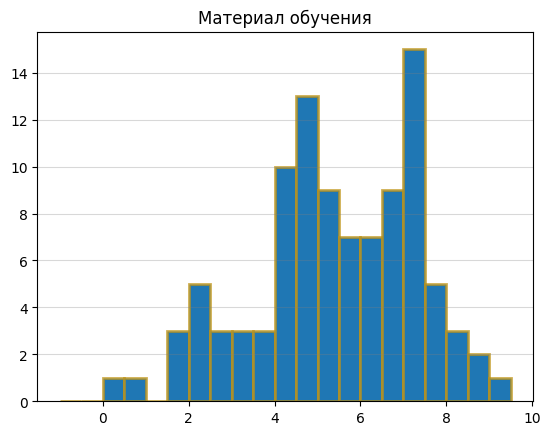
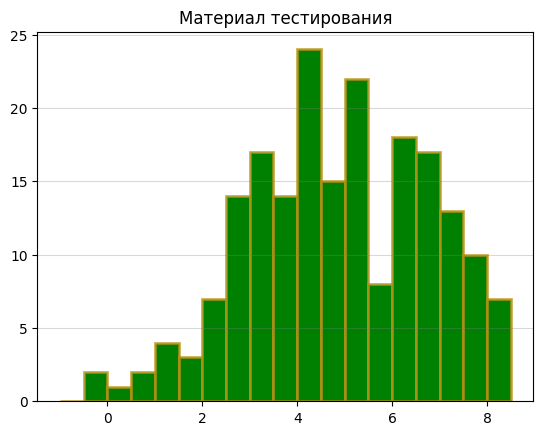
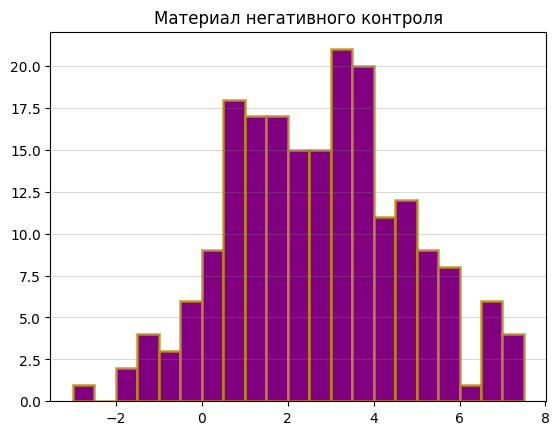
Исходя из графиков, я выбрала порог веса 4, так как в этой точке на гистограмме для материала обучения количество последовательностей с данным весом резко начинает расти, а на гистограмме для отрицательного контроля – резко падать. Там же наблюдается пик на гистограмме для материала тестирования. В области веса 6 наблюдается какой-то странный провал, как будто что-то пошло не так, так что на это место я не обращала внимания.
Результат проверки представлен в таблице 2:
| Обучающая выборка | Тестовая выборка | Отрицательный контроль | |
|---|---|---|---|
| + сигнал | 81 (81,0%) | 136 (68,0%) | 52 (26,0%) |
| - сигнал | 19 (19,0%) | 64 (32,0%) | 148 (74,0%) |
Вычисление IC и постороение LOGO
Вычисленное информационное содержание выравнивания:
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| A | -0,02 | -0,09 | -0,11 | -0,11 | 0,29 | -0,02 | -0,06 | 1,76 | 0,00 | 0,00 | -0,08 | -0,02 | -0,14 |
| T | -0,11 | -0,10 | -0,15 | -0,12 | -0,08 | -0,14 | -0,14 | 0,00 | 1,76 | 0,00 | -0,16 | -0,16 | -0,12 |
| G | 0,05 | 0,36 | 0,09 | 0,02 | 0,32 | 0,01 | 0,18 | 0,00 | 0,00 | 2,29 | 0,75 | 0,04 | 0,46 |
| C | 0,13 | -0,03 | 0,36 | 0,36 | -0,07 | 0,25 | 0,34 | 0,00 | 0,00 | 0,00 | -0,09 | 0,36 | 0,01 |
| IC(j) | 0,05 | 0,13 | 0,19 | 0,15 | 0,46 | 0,10 | 0,33 | 1,76 | 1,76 | 2,29 | 0,43 | 0,22 | 0,21 |
Построила LOGO для тестовой выборки:
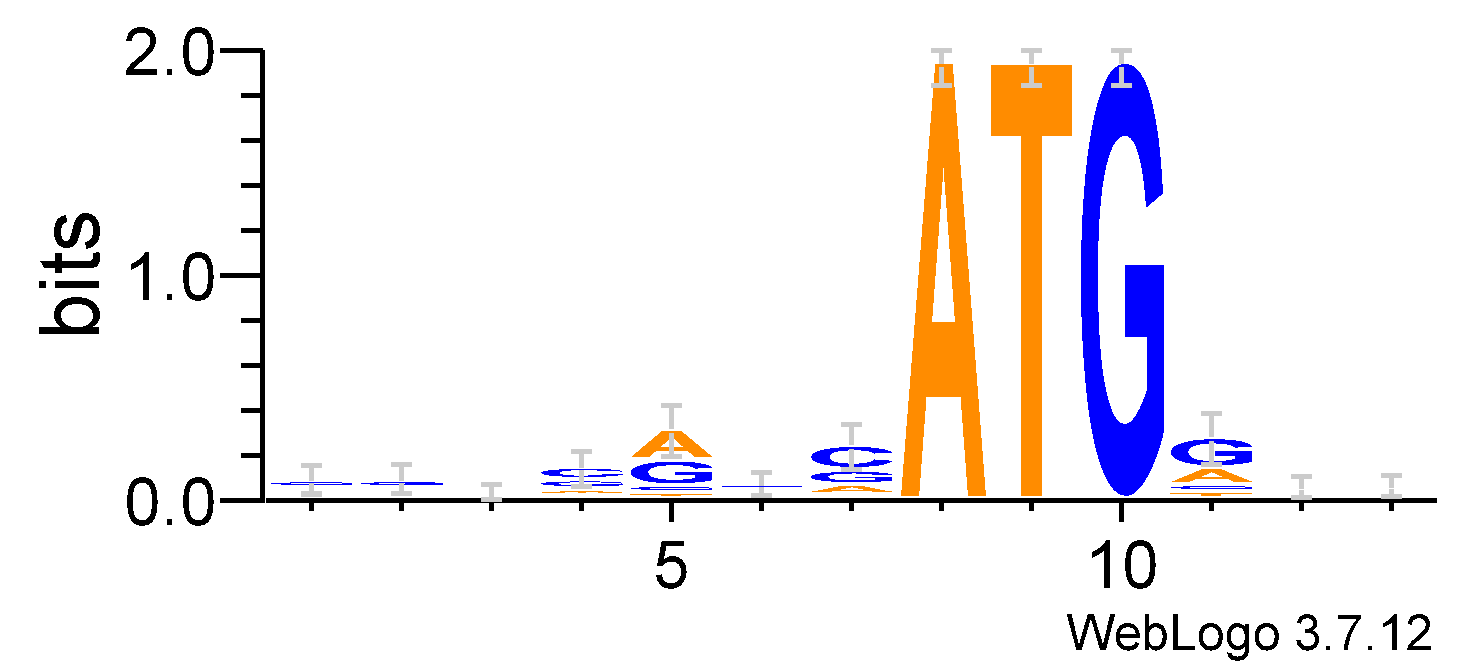
Очень хорошо видно консервативные ATG, а также наиболее заметны позиции вокруг ATG: 5, 7 и 11.