Практикум 3. Укоренение. Сравнение деревьев. Бутстрэп
1. Реконструкция дерева по нуклеотидным последовательностям
Для выбранных животных реконструируем дерево по последовательностям малой РНК митохондриальных рибосом (12S rRNA).
Митходриальный геном был найден не для всех видов, поэтому для этих организмов были взяты ближайшие к ним виды:
1. Для Microtus arvalis был взят Microtus obscurus. Мнемоника: 9RODE
2. Для Phodopus campbelli - Phodopus sungorus. Мнемоника: PHOSU
3. Для Hystrix africaeaustralis - Hystrix indica. Мнемоника: 9HYST
4. Для Thomomys monticola ближайщий родственник Thomomys bottae, но в EMBL файле для него не было координат для 12S rRNA, только join(JAVLKH010000001.1:1..17079), поэтому был взят родственник из другого рода Geomys pinetis. Мнемоника: GEOPI
5. Для Aplodontia rufa тоже не нашлись родственники того же рода, поэтому для построения дерева был взят Sciurus vulgaris. Мнемоника: SCIVU
Последовательности были выровнены и записаны в файл 12S_rRNA_alignment.fasta. Потом перезаписали файл в формат phy и реконструировали дерево программой IQ-Tree.

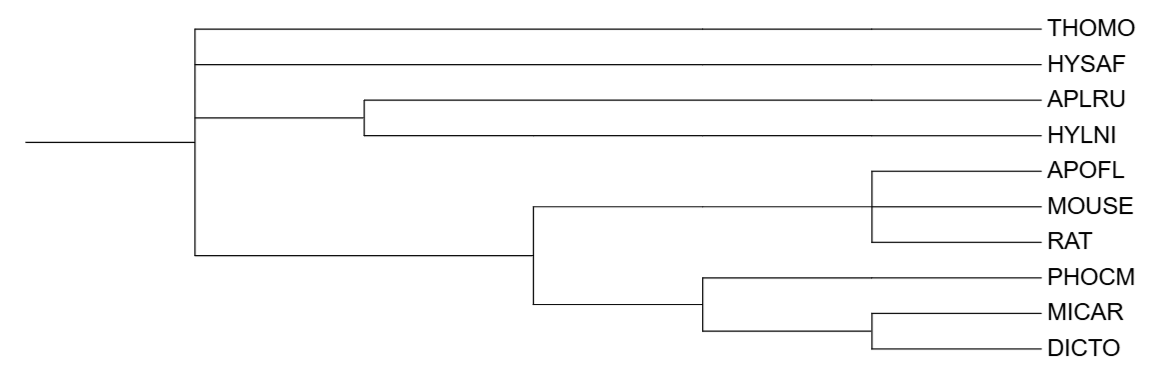
Сравнение дерева 12S rRNA с деревом видов: на обоих деревьях четко выделяются общие кластеры, следовательно 12S rRNA удобно использовать для разрешения родства на уровне семейств:
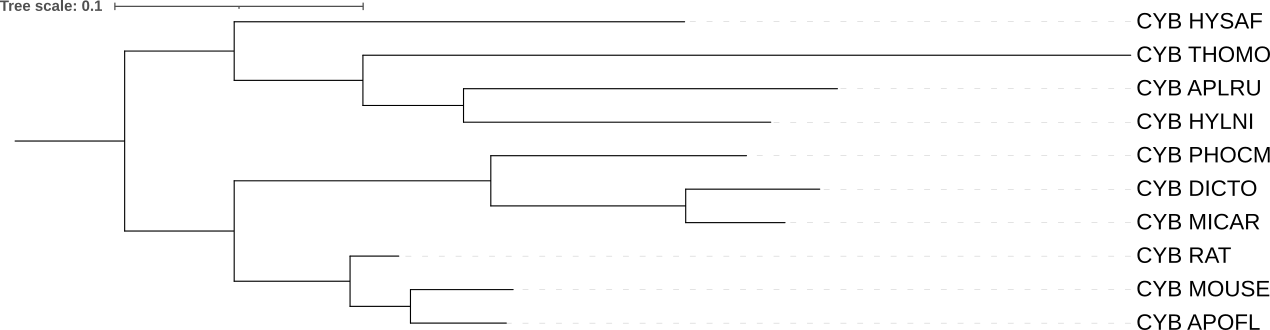
Сравнение дерева 12S rRNA с деревом цитохромов В:
На дереве 12S rRNA: видно, что дикобраз (9HYST) и гребнепалая мышь (HYLNI) объединяются с белкой (SCIVU) в один крупный кластер. При этом гофер (GEOPI-THOMO) вынесен на самую дальнюю ветвь.
PHOSU (хомячок) и DICTO/9RODE (полевки) сгруппированы очень тесно.
Ветви выглядят более сбалансированными по длине, что характерно для рибосомальных генов, которые эволюционируют с более строгими структурными ограничениями.
Дерево CytB: Топология меняется. Здесь дикобраз (HYSAF) и гофер (THOMO) оказываются в одном кластере в верхней части дерева, отделенном от остальных грызунов. Это дерево ломает монофилию подотряда Hystricomorpha (дикобразообразных).
Триада RAT, MOUSE, APOFL (лесные мыши) разрешена очень четко с короткими терминальными ветвями. Цитохром b, будучи белок-кодирующим геном, часто лучше разрешает отношения внутри семейств из-за высокой вариабельности в третьих позициях кодонов.
Ветвь к THOMO (гоферу) выглядит слишком длинной (это может быть связано с тем, что последовательность цитохрома b у этого таксона накопила критическое количество замен).
2. Укоренение во внешнюю группу
Все текущие животные (мыши, крысы, хомяки, белки, гоферы, дикобразы) относятся к отряду Rodentia (Грызуны). Внешней группой для грызунов является Зайцеобразные (Lagomorpha). Они являются ближайшими родственниками грызунов (вместе они составляют надотряд Glires), но при этом они эволюционно отделились от них достаточно давно. В качестве внешней группы возьмём заяца-русака (Lepus europaeus) - AJ421471.

На первом дереве (Outgroup): гофер (GEOPI-THOMO) расположен глубже внутри дерева и ближе к кластеру мышеобразных (Muroidea),
а дикобразы и белки образовали свои отдельные базальные ветви.
Заяц (Lepus) занял позицию самой дальней ветви, так как зайцеобразные отделились от общего ствола с грызунами еще до того, как сами грызуны разделились на подотряды.
Внешняя группа расставила грызунов в более естественном порядке.
Базальные грызуны: первыми после зайца отходят ветви, ведущие к дикобразам (9HYST) и белкам/гребнепалым (SCIVU/HYLNI). Эта топология соответствует классическим представлениям о разделении подотрядов Hystricomorpha и Sciuromorpha.
Укоренение в среднюю точку — это чисто математический метод, который предполагает, что скорость эволюции во всех ветвях одинакова. На втором дереве (Midpoint) ветвь GEOPI-THOMO выглядит как сестринская группа ко всем остальным грызунам. Поскольку у гофера эта ветвь очень длинная,
алгоритм ошибочно принял её за самую древнюю. Это создаёт ложное впечатление, что эта группа отделилась раньше всех.
Монофилия Muroidea: Мыши, крысы, хомяки и полевки по-прежнему образуют единый, компактный и хорошо выраженный кластер.
Укоренение во внешнюю группу выполнено верно. Оно исправило главную ошибку визуализации — искусственное выделение гофера в качестве самой древней группы.
Теперь дерево отображает не просто математические расстояния между последовательностями, а иерархию происхождения от общего предка.
3. Бутстреп
Теперь повторим филогенетическую реконструкцию программой fastme, использовав 100 реплик бутстрепа.

При сравнении реконструированного дерева со 100 репликами бутстрепа с деревом видов так же видно, что основная структура сохранилась, но есть локальные изменения внутри групп. Группа APLRU + HYLNI (беличьи) реконструирована идеально с поддержкой 100. Разделение на две большие ветви (мышиные и хомяковые) также подтверждено поддержкой 100. В группе хомяковых топология (PHOCM внешняя к MICAR + DICTO) совпадает с деревом видов, но здесь поддержка 50 и это самый слабый узел на дереве. Хотя ветвь и совпала с эталоном, FastME не уверен в этом результате на данных для 12S rRNA. Узел (APOFL + RAT) - поддержка 78; на дереве видов этот узел неразрешен. Поддержка 78 считается умеренной, тем не менее в 22% реплик бутстрепа программа видела другую картину (скорее всего ту, где крыса с мышью вместе).