Практикум 4. Реконструкции филогении
В данном практикуме необходимо было найти в протеомах отобранных 7 бактерий достоверные гомологи белка CLPX_ECOLI (ATP-связывающую субъединицу ATP-зависимой Clp протеазы Escherichia coli) с помощью программы blastp с поогом на E-value 0,0001.
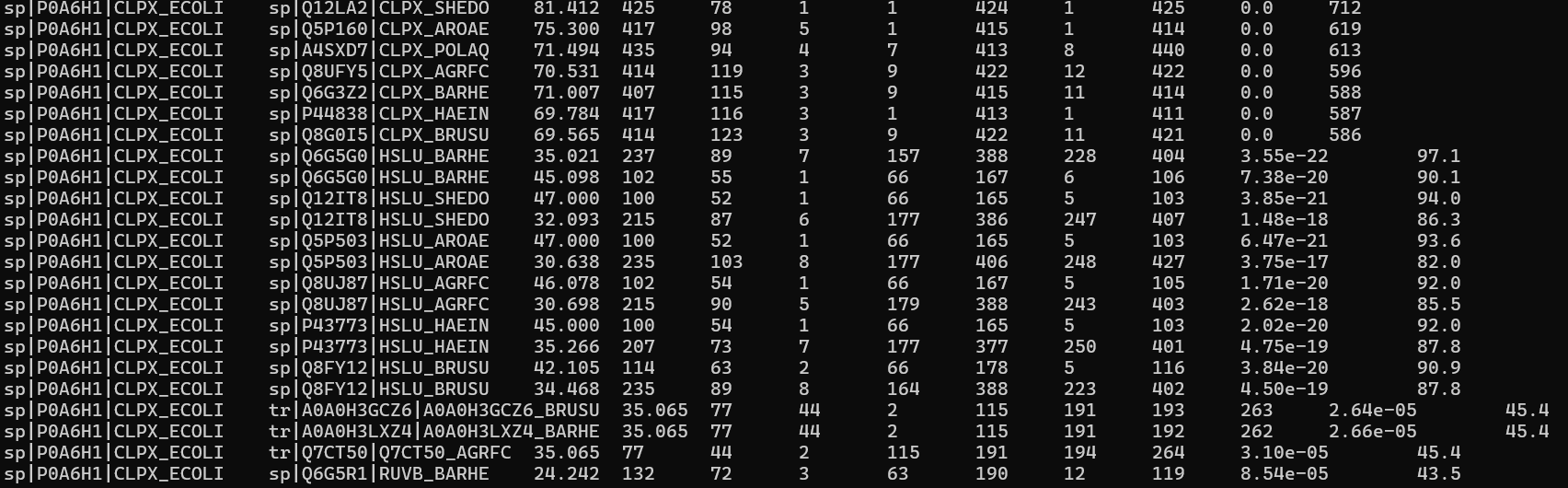
Далее для отобранных последовательностей с помощью программы fastme с эволюционной моделью MtREV и 100 репликами бутстрепа было реконструировано филогенетическое дерево.

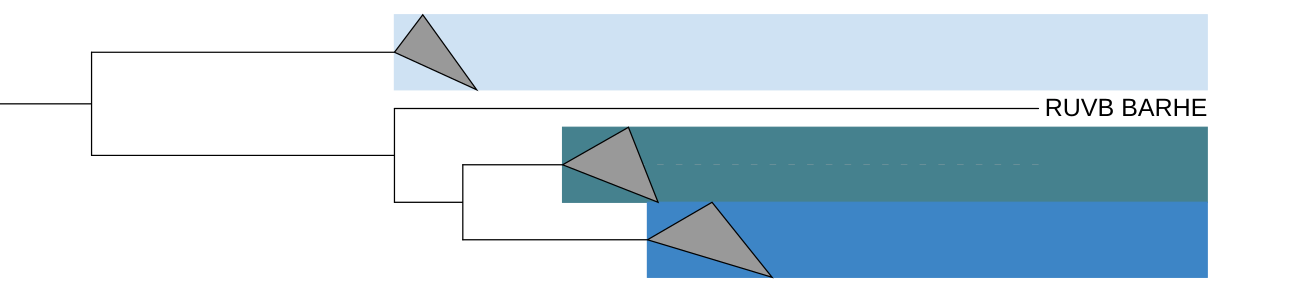
Три пары паралогов: белки из одного организма, появившиеся в результате дупликации гена в прошлом
1. CLPX_BARHE и HSLU_BARHE
2. CLPX_AGRFC и HSLU_AGRFC
3. CLPX_BRUSU и HSLU_BRUSU
Также паралогами будут, например, CLPX_BARHE и RUVB_BARHE, так как они гомологичны, но выполняют разные функции в одной клетке.
Три пары ортологов: один и тот же белок в разных организмах, разделившийся в момент расхождения видов
1. CLPX_BARHE и CLPX_BRUSU
2. HSLU_HAEIN и HSLU_SHEDO
3. CLPX_HAEIN и CLPX_SHEDO
Видно, что дерево сначала делится на функциональные группы (ClpX отдельно, HslU отдельно). Это говорит о том, что дупликация, создавшая эти два семейства, произошла гораздо раньше, чем разделились виды бактерий. Группу RUVB_BARHE можно охарактеризовать как высокодивергентного удаленного гомолога. Хотя RuvB, ClpX и HslU относятся к одному суперсемейству АТФаз, их функции в клетке кардинально различаются:
ClpX и HslU — это шапероны/анфолдазы, которые участвуют в деградации белков
RuvB — это ДНК-хеликаза, участвующая в рекомбинации и репарации ДНК.
Сравнение реконструированной филогении ортологических групп с филогенией бактерий.
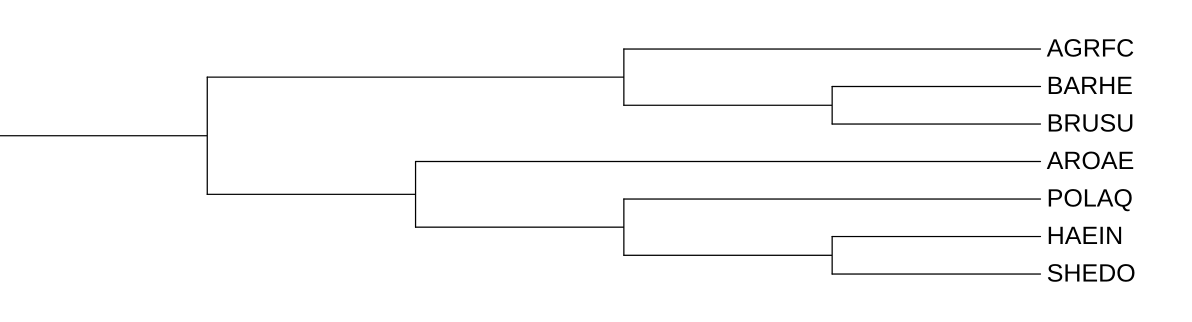
1. Группа HslU:
В группе отсутствуют белки только для POLAQ.
Несмотря на отсутствие одного вида, иерархия сохраняется: пара Гамма-протеобактерий (HAEIN, SHEDO) отделена от остальных.
Альфа-протеобактерии (AGRFC, BARHE, BRUSU) образуют монофилетическую ветвь с правильной внутренней структурой (BRUSU и BARHE ближе друг к другу).
Но также наблюдается топологическое несоответствие с филогенией бактерий: представитель Бета-протеобактерий (AROAE) образует кластер с Гамма-протеобактериями, в то время как согласно таксономии он должен быть ближе к Альфа-протеобактериям.
2. Группа ClpX:
Представлена белками изо всех 7 исследуемых бактерий.
Белки Гамма-протеобактерий (HAEIN, SHEDO) не образуют обособленную (монофилетическую) ветвь по отношению к остальным. Вместо этого наблюдаем лестничную структуру. Сначала отделяется ветвь, ведущая к HAEIN. Затем отделяется ветвь к SHEDO. Ниже по дереву формируется общий кластер Бета- и Альфа-протеобактерий. Это парафилия (когда группа включает общего предка, но не всех его потомков в рамках одной ветви). Возможно если белок ClpX у HAEIN эволюционировал значительно быстрее или медленнее, чем у SHEDO, алгоритм мог не объединить их в одну пару.
Бета-протеобактерии (AROAE, POLAQ) группируются вместе.
Альфа-протеобактерии образуют кластер, где AGRFC и BRUSU наиболее близки, а BARHE примыкает к ним. Это ошибка реконструкции. Согласно дереву видов, BARHE и BRUSU должны быть наиболее близки друг к другу, а AGRFC — быть для них внешней группой.
3. Верхний кластер:
Внутри этого небольшого кластера видим, что AGRFC и BRUSU оказались ближе друг к другу, а BARHE — дальше. Опять-таки на дереве таксономии ближе друг к другу BRUSU и BARHE.