Практикум 7. Трансмембранные белки
Трансмембранный α-спиральный белок: 4kyt - Кальциевая АТФаза (Calcium ATPase, E2 state (Ca-free), complex with phospholamban)
UniProt: P04191
Организм: Oryctolagus cuniculus (Дикий кролик)
Расположение: мембрана ЭПР
Функция: ключевой регулятор работы поперечнополосатых мышц, выступающий в качестве основной Ca2+-АТФазы, ответственной за обратный захват цитозольного Ca2+ в саркоплазматический ретикулум. Катализирует гидролиз АТФ, сопровождающийся перемещением кальция из цитозоля в просвет саркоплазматического ретикулума
Координаты трансмембранных участков белка: А (сама кальциевая АТФаза): 1( 60- 78), 2( 86- 105), 3( 260- 279), 4( 291- 306), 5( 760- 781), 6( 789- 807), 7( 832- 853), 8( 896- 916), 9( 933- 950),10( 967- 987) В и С регуляторные белки (фосфоламбан), которые образуют комплекс с АТФазой B: 1( 28- 48) C: 1( 27- 38)
Запустим DeepTMHMM для последовательности этого белка, взятой из PDB, чтобы предсказать трансмембранные участки
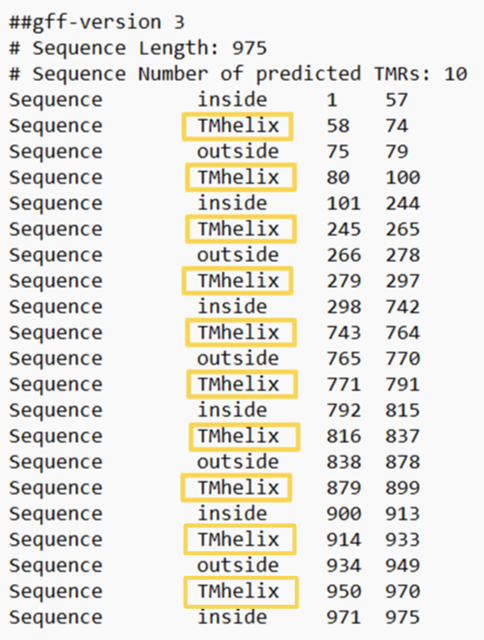
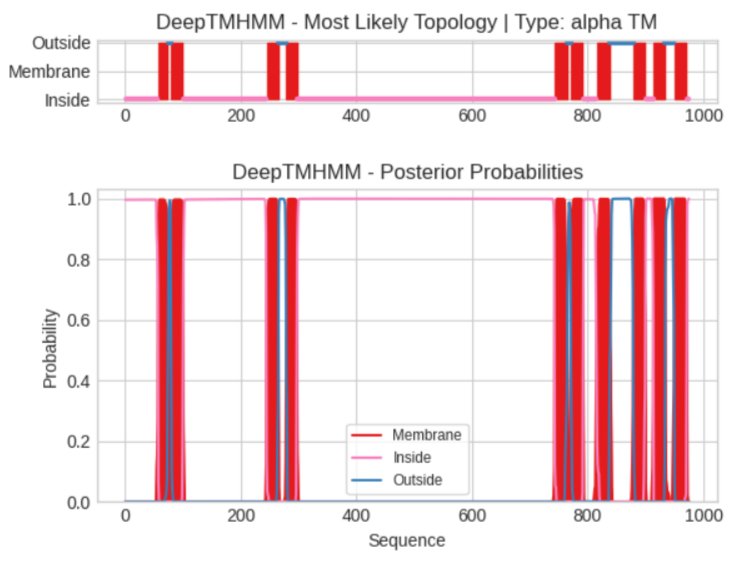
На рисунке 2 верхний график Most Likely Topology - анализирует всю последовательность и выдает четкую схему расположения белка, красные блоки - трансмембранные спирали (10 штук совпадает с данными с OPM), розовая линия - участки белка, которые находятся внутри, синяя линия - участки белка, находящиеся с внешней стороны.
Верхний график — это вердикт программы, а нижний — это предположения программы. По оси Y здесь отложена вероятность от 0.0 до 1.0. Красные пики: показывают вероятность того, что конкретный аминокислотный остаток принадлежит мембране. Если пик узкий и доходит до единицы, программа абсолютно уверена в наличии спирали. Розовая и синяя линии: показывают вероятность того, что остаток находится внутри или снаружи.
Для 9 трансмембранных участков предсказания перекрываются:
1) OPM (60-78) и DeepTMHMM (58-74) перекрытие на 15 аминокислот.
2) OPM (86-105) и DeepTMHMM (80-100) перекрытие на 15 аминокислот.
3) OPM (260-279) и DeepTMHMM (245-265) перекрытие на 6 аминокислот.
4) OPM (291-306) и DeepTMHMM (279-297) перекрытие на 7 аминокислот.
5) OPM (760-781) и DeepTMHMM (743-764) перекрытие на 5 аминокислот.
6) OPM (789-807) и DeepTMHMM (771-791) перекрытие на 3 аминокислоты.
7) OPM (832-853) и DeepTMHMM (816-837) перекрытие на 6 аминокислот.
8) OPM (896-916) и DeepTMHMM (879-899) перекрытие на 4 аминокислоты.
9) OPM (933-950) и DeepTMHMM (914-933) не перекрываются (участок (933-950) не замечен в DeepTMHMM).
10) OPM (967-987) и DeepTMHMM (950-970) перекрытие на 4 аминокислоты.
Результаты, полученные двумя методами, сильно различаются. Перекрытия для трансмембранных участков слишком короткие.
Причины различия результатов:
OPM: Основан на реальных 3D-структурах (из PDB), полученных методом рентгеноструктурного анализа или криоэлектронной микроскопии. Границы ТМ-участков здесь определяются по термодинамике взаимодействия аминокислот с липидным бислоем.
DeepTMHMM: Это математическая модель, которая анализирует только последовательность. Она «угадывает» положение спиралей на основе статистических закономерностей и обучения нейросети на тысячах других белков. Она не видит реальных физических взаимодействий в конкретном кристалле белка. DeepTMHMM анализирует локальную гидрофобность. Если спираль находится в центре белка и окружена другими спиралями, а не липидами напрямую, её гидрофобный профиль может быть смазан (спирали собраны в плотный пучок, внешние спирали напрямую контактируют с липидами мембраны. Они обычно очень гидрофобны, и программа легко их находит, а внутренние спирали находятся в центре пучка и окружены другими спиралями. Поскольку они контактируют с белком, а не с жиром, в их составе могут появиться полярные или заряженные аминокислоты). Это часто приводит к тому, что программа сдвигает границы спирали или вообще теряет сегмент (как в пункте 9), если он недостаточно выражен по гидрофобности.
База данных TCDB
3 (Класс): Первично-активные транспортеры - насос
A (Подкласс): Гидролазы пирофосфатных связей
3 (Семейство): АТФазы P-типа
2 (Подсемейство): Ca2+-транспортирующих АТФаз
43 (Система): Конкретный представитель — Фосфоламбан - это регуляторный белок, который «тормозит» работу кальциевого насоса.
1 (Кдасс): пассивные каналы - не тратят энергию АТФ, а просто позволяют молекулам проходить сквозь мембрану по градиенту концентрации
B (Подкласс): порины, бета-бочонок
21 (Семейство): внешних мембранных белков G - порины, которые служат общим каналом для прохода различных сахаров и других питательных веществ
1.1 (Система): указывает на конкретный белок — OmpG из Escherichia coli