Аннотация
Проведён биоинформатический анализ генома почвенной бактерии Adhaeribacter radiodurans. Установлено распределение межгенных промежутков генома (медиана - 250 п.н.). Мода GC состава кодирующих участков - 43-44%, что является нехарактерным показателем для радиорезистентных бактерий. Распределение генов резко неоднородно: выявлены «пустые» участки (6.4% генома) и сверхплотные кластеры (5.7%). Протеом типичен для бактерий (медиана длины белка 288 а.о.), но включает экстремально длинные белки. Полученные данные в ходе данного мини-обзора скорее указывают на адаптацию к ризосфере, а не на классическую радиорезистентность.
Введение
Adhaeribacter radiodurans – вид граммотрицательных бактерий, принадлежащий к семейству Hymenobacteraceae в филуме Bacteroidota [1]. Данный микроорганизм был выделен из ризосферы (слоя почвы, прилегающего к корням растения, толщиной не более 2–5 мм) [3] злака Elymus tsukushiensis, растущего на островах Докдо в Республике Корея. Клетки этой бактерии представляют собой неподвижные палочки размером от 0,7 до 1,0 мкм в ширину и от 2,5 до 5,5 мкм в длину. У бактерии данного вида присутствуют такие ферменты как каталаза (катализирует разложение пероксида водорода) и оксидаза (участвует в катаболизме) [1]. Предположительно, это может свидетельствовать о аэробности или микроаэрофильности бактерии. Видовое название «radiodurans» было дано из-за предполагаемой радиационной устойчивости организма к излучению. Однако для подтверждения его радиационной резистентности и выяснения других физиологических особенностей необходимы дальнейшие исследования [2]. В рамках данного мини-обзора были построены гистограммы длин белков, межгенных промежутков, в том числе пересекающихся, исследовано распределение CDS по GC-составу, а также плотность генома.
Таблица 1. Таблица полной таксономии Adhaeribacter radiodurans [4]
| Domain | Bacteria |
|---|---|
| Kingdom | Pseudomonadati |
| Phylum | Bacteroidota |
| Class | Cytophagia |
| Order | Cytophagales |
| Family | Hymenobacteraceae |
| Genus | Adhaeribacter |
| Species | Adhaeribacter radiodurans |
Методы
Аннотированный геном Adhaeribacter radiodurans был скачан с сайта NCBI с помощью команды wget в командной оболочке Bash. Для подсчета количества значений, попадающих в заданный карман для дальнейшего построения гистограммы была использована команда =COUNTIFS. Для подсчета межгенных промежутков координаты CDS были отсортированы по возрастанию и была задана формула вычитающая координату начала следующего кодирующего участка из предыдущего. Статистические показатели были вычислены с помощью соответствующих формул: =STDEV.P (стандартное отклонение), =MIN (минимальное значение), =MAX (максимальное значение), =AVERAGE (среднее значение), =MODE (мода) В ходе исследования плотности генома для определения, к какому окну принадлежит кодирующий участок использовалась формула = CEILING. Все задания данного мини-обзора были выполнены с помощью инструментария электронных таблиц Google Sheets (построение графиков, диаграмм, вычисление статистических показателей) и командной оболочки Bash (работа с файлами)
Результаты
Длины белков, закодированных в геноме Adhaeribacter radiodurans
Для протеома бактерии было определено распределение белков по длине, составлена гистограмма (Рисунок 1) и подсчитаны некоторые статистические показатели (Таблица 2), а также составлена гистограмма белков с наибольшей длиной (Рисунок 2). Для более подробного ознакомления см. ссылку S2 сопроводительных материалов.
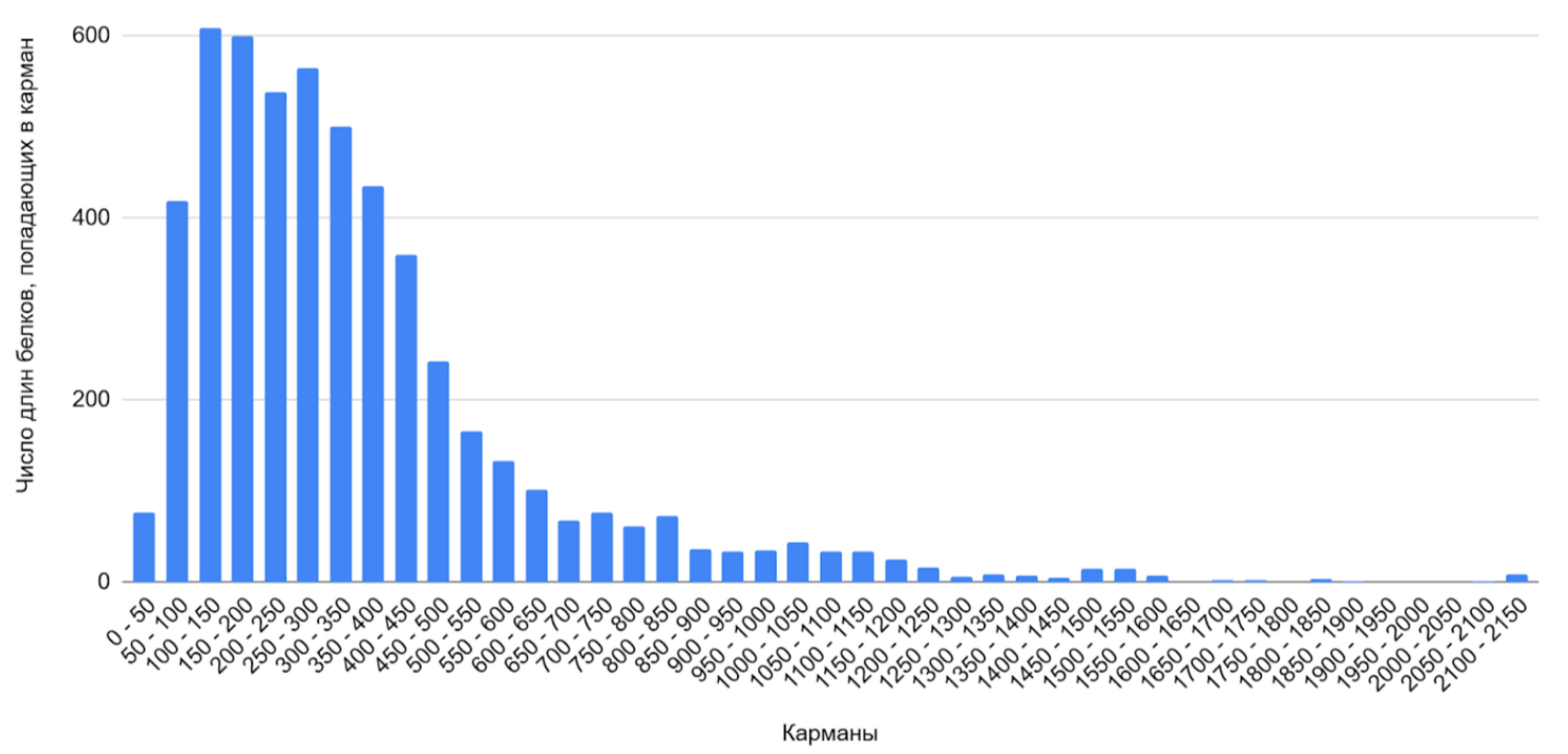
Таблица 2. Статистические показатели распределения длин белков (аминокислот)
| Статистический показатель | Значение |
|---|---|
| Максимальная длина белка | 5441 |
| Минимальная длина белка | 23 |
| Медианная длина белка | 288 |
| Среднее значение длины белка | 356.6 |
| Стандартное отклонение длины белка | 289.5 |
Основная масса белков — короткие (до 350 а.о.), что типично для бактериальных протеомов. Длинные белки (свыше 2400 а.о.) — исключение, но они есть, что может быть связано с наличием белковых комплексов, особенностями метаболизма или адаптации к условиям среды (в том числе радиорезистентности)
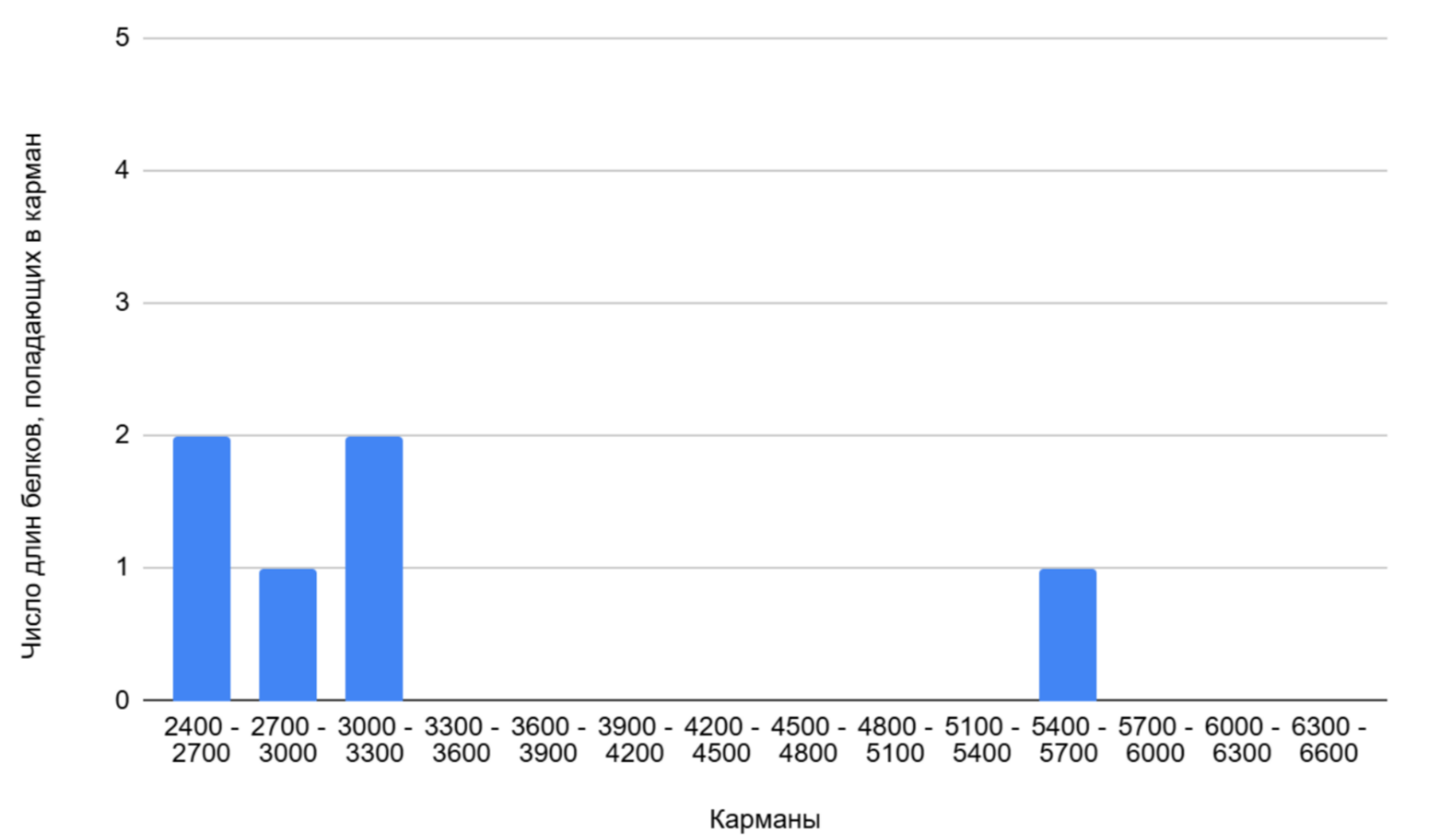
На второй гистограмме изображен «хвост» распределения (Рисунок 2), показывающий белки, имеющие наибольшие показатели длины. Интервалы шире, чем в первой гистограмме, что позволяет лучше визуализировать редкие случаи. Больше всего белков с длинами, попадающими в карманы: 2400-2700 и 3000-3300 (по 2). Всего диаграмме представлены 6 самых длинных белков бактерии (Таблица 3):
Таблица 3. Сравнительная таблица самых длинных белков в геноме
| Название | Длина продукта |
|---|---|
| malectin domain-containing carbohydrate-binding protein | 5441 |
| PKD domain-containing protein | 3230 |
| malectin domain-containing carbohydrate-binding protein | 3217 |
| LamG-like jellyroll fold domain-containing protein | 2774 |
| isopeptide-forming domain-containing fimbrial protein | 2616 |
| cell surface protein SprA | 2434 |
1) Malectin domain-containing carbohydrate-binding protein - углевод-связывающий белок, содержащий малектиновый домен, является сенсорной структурой в системе контроля качества гликопротеинов: специализируется на распознавании определенного типа дефектных или неправильно свернутых гликопротеинов [7]
2) PKD domain-containing protein — это «датчик потока и состава среды» в бактерии. Располагается на поверхности, часто на ресничке [6]
3) LamG-like jellyroll fold domain-containing protein - располагается на внешней поверхности клетки или в межклеточном пространстве. Специализируется на мультивалентных взаимодействиях - способен надёжно, специфично и часто многократно соединять разные биологические "детали" (клетки, белки, компоненты матрикса) в единую, стабильную и функциональную трехмерную структуру [9]
4) Isopeptide-forming domain-containing fimbrial protein - строительный блок (субъединица) пили, обеспечивает ее экстремальную механическую прочность посредством изопептидной связи. Изопептидная связь — ковалентная связь между боковыми группами аминокислот (чаще всего лизина и аспарагина/глутамина), расположенных внутри одного белка
5) Cell surface protein SprA - Поверхностный белок, ответственный за выброс токсичных белков через трансмембранный канал (пору), при этом это часто предполагает гибель самой клетки (автолиз) [10]
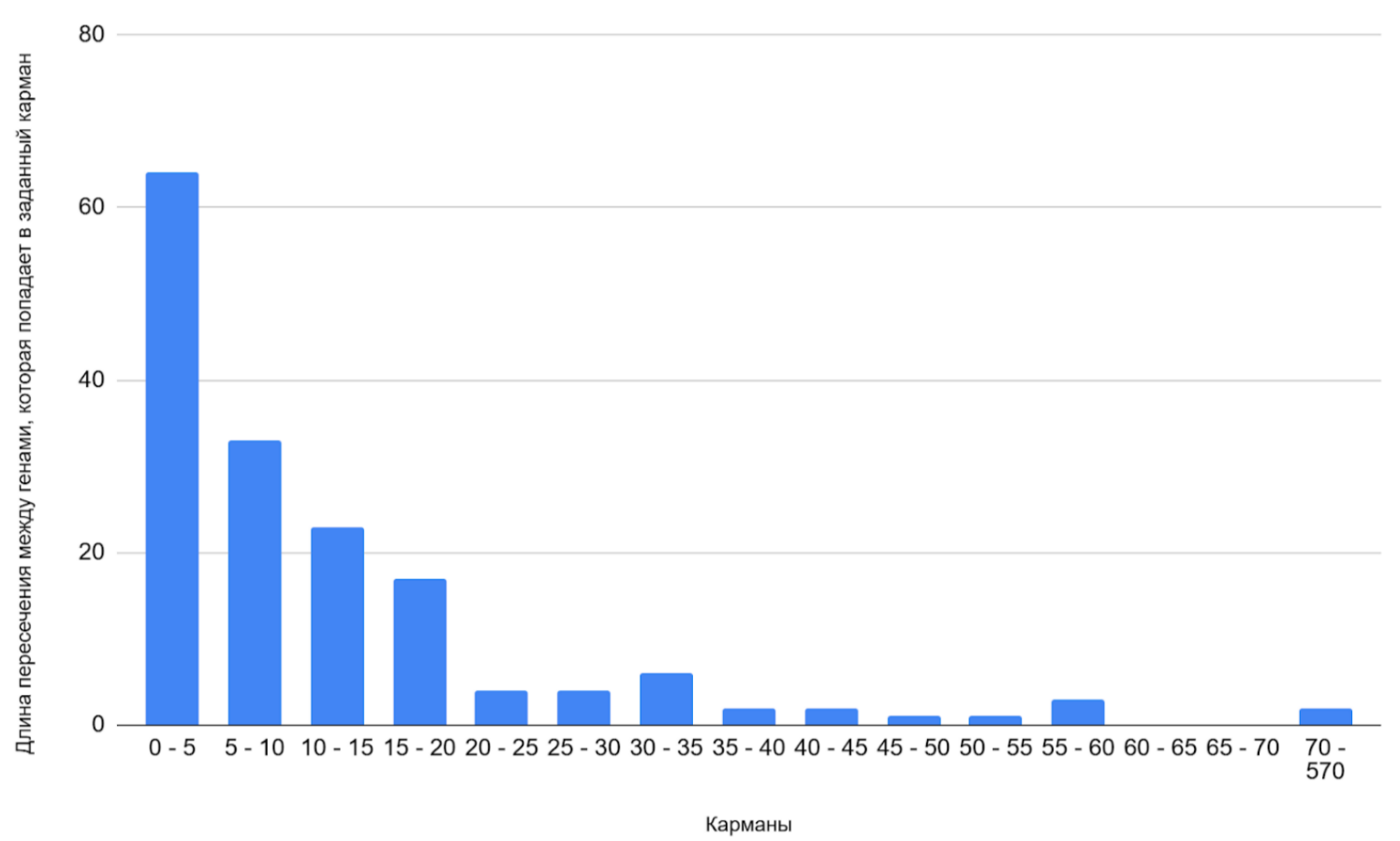
Межгенные промежутки в геноме Adhaeribacter radiodurans
Для генома была построена гистограмма межгенных промежутков (см. ссылку S1 сопроводительных материалов) с карманом в 20 пар оснований (Рисунок 3). Расстояние, равное нулю соответствует расположенным вплотную генам, расстояние, меньше нуля - пересекающимся генам. Гистограмма межгенных промежутков в геноме Adhaeribacter radiodurans демонстрирует унимодальное распределение (единственный пик). Ярко выраженный пик приходится на область коротких промежутков между генами - 0- 20 п.н., что может говорить о возможной оперной активности генома исследуемой бактерии.
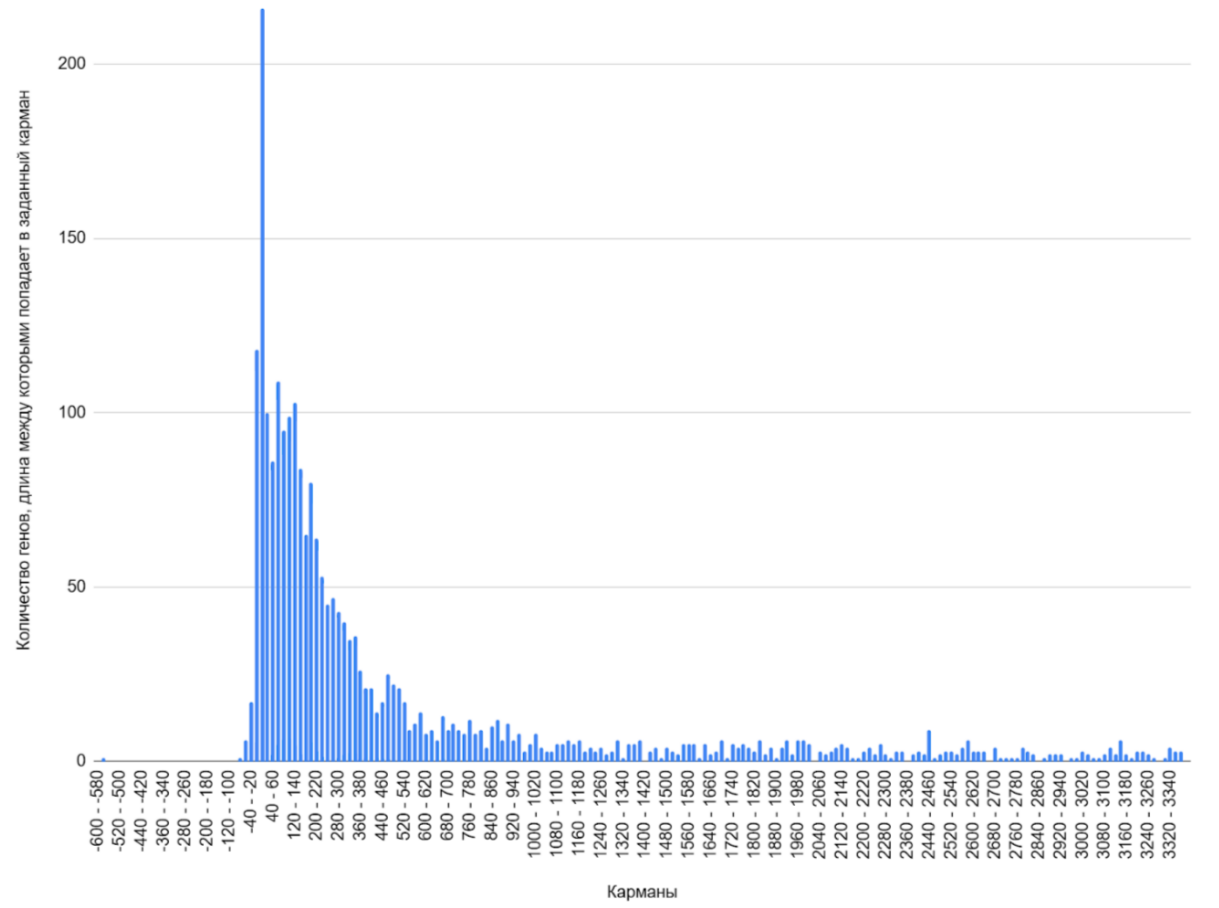
Таблица 4. Статистические показатели распределения длин межгенных промежутков (пар оснований)
| Параметр | Плюс цепь | Минус цепь |
|---|---|---|
| Максимальная длина | 39068 | 39542 |
| Минимальная длина | -566 | -6796132 |
| Медианная длина | 250 | 246 |
| Мода межгенного промежутка | -3 | -3 |
Для сравнительного анализа был проанализирован генетический материал CDS на минус цепи (Рисунок 5). Единственное значительное отличие - значение самого короткого промежутка: -6796132 пар оснований (Таблица 4)
Также отдельно была построена гистограмма пересечений между генами (Рисунок 6). Пики гистограммы указывают на области сверхвысокой плотности генов: наибольшее количество перекрывающихся кодирующих областей приходятся на карман 0-5 п.н.
Распределение CDS бактерии Adhaeribacter radiodurans по GC-составу
Содержание гуанина и цитозина в геноме составляет 40,8 моль% [1]. Гистограмма показывает унимодальное распределение (см. таблицу S2 сопроводительных материалов) с пиком в области 43-44%, что соответствует низкому общему GC-составу генома данной почвенной бактерии (Рисунок 7). Распределение имеет слабую положительную асимметрию (длинный хвост немного перемещен в сторону высоких GC).
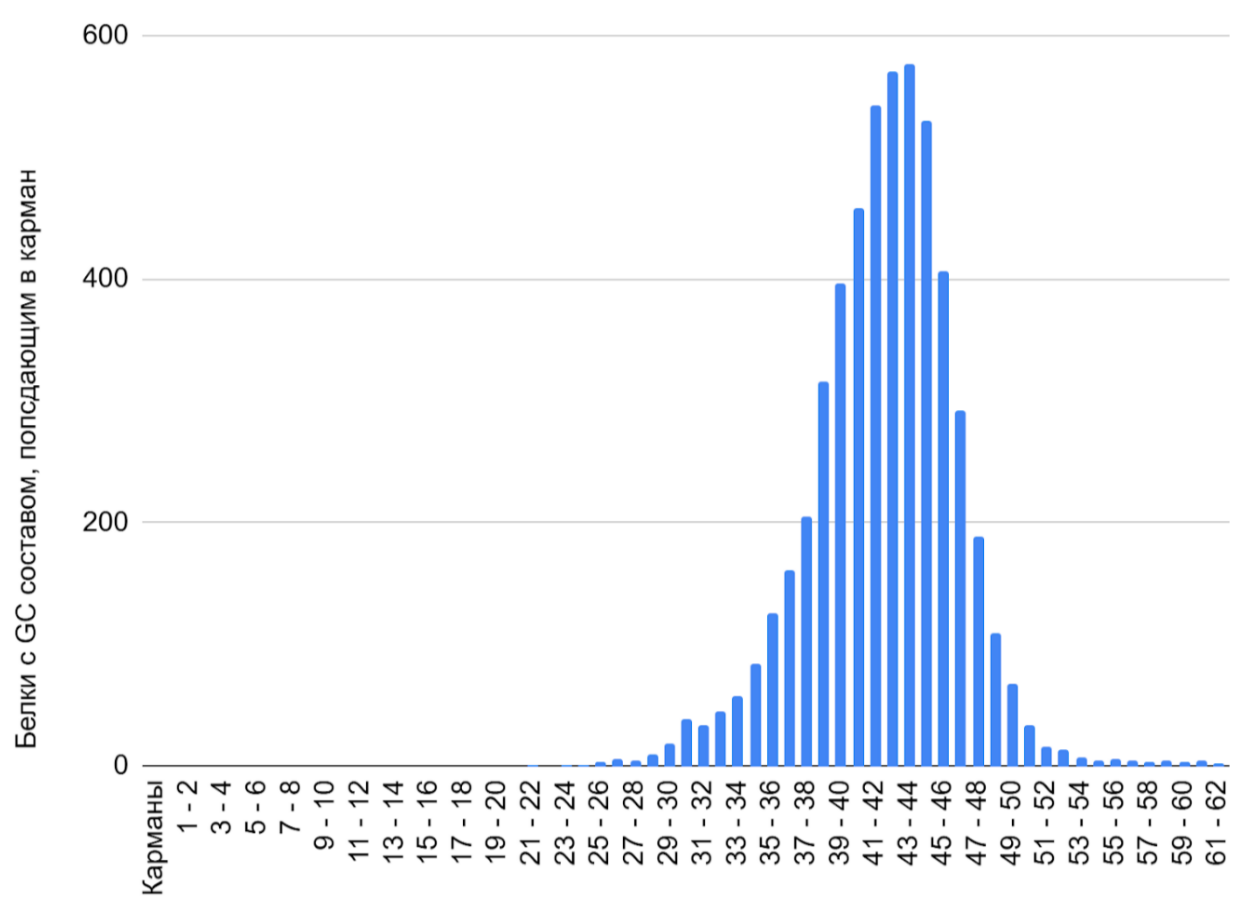
Пограничное значение GC состава (между средним и низким) указывает на некоторое противоречие: высокий GC состав обычно встречается у экстремофилов (термофилов или радиоустойчивых организмов), так как высокие температуры и повреждающие факторы требуют стабильной ДНК, которую обеспечивает высокий GC [11]. Однако видовое название radiodurans было дано именно из-за предполагаемой радиорезистентности, но, если ограничиться исключительно GC составом, из гистограммы Adhaeribacter radiodurans можно сделать вывод, что она таковым не является. Для микроорганизмов с низким GC составом характерна ассоциация с организмами (комменсализм, паразитизм, симбиоз) или анаэробными условиями обитания [12], что подтверждается обитанием этой бактерии в ризосфере.
Плотность генома Adhaeribacter radiodurans
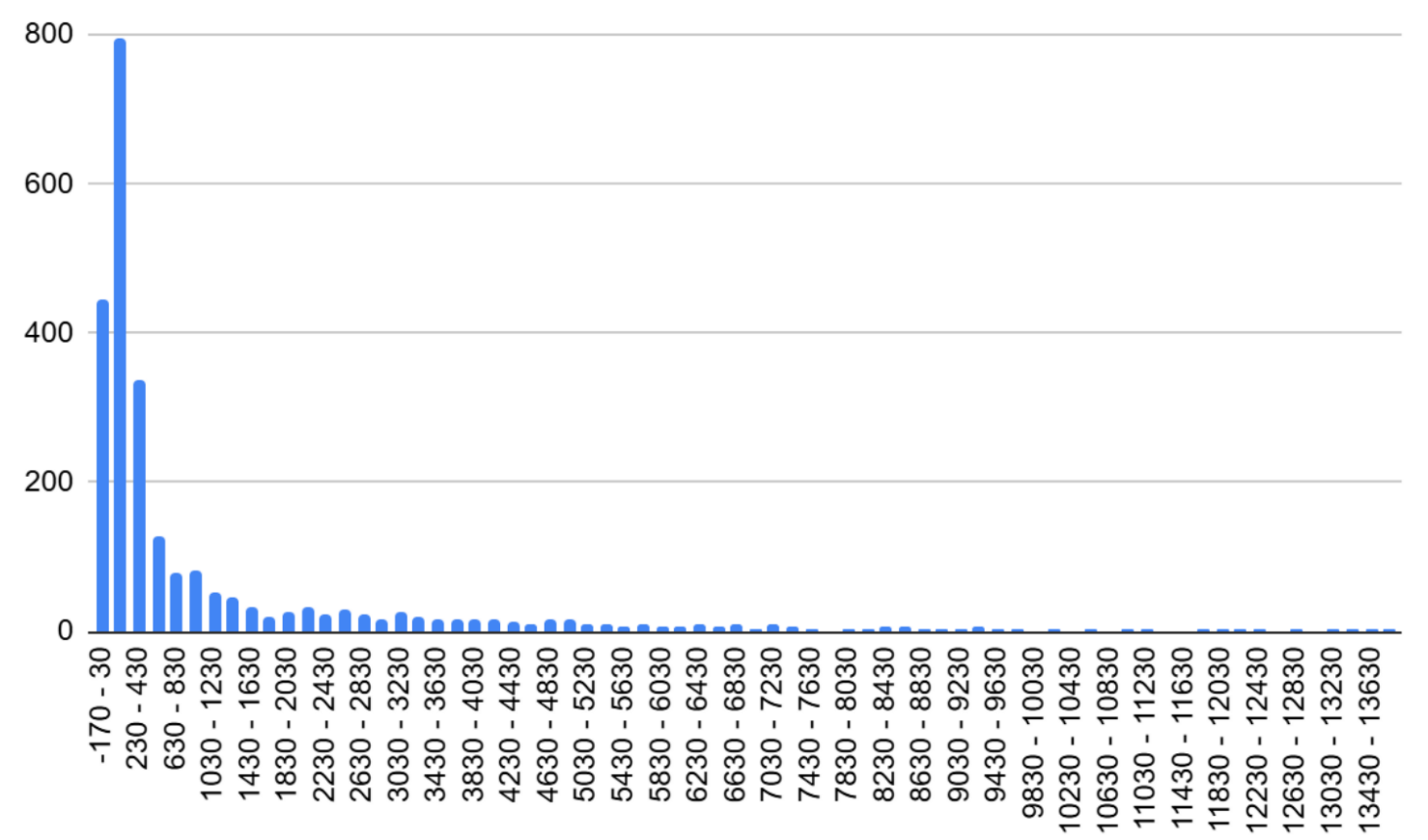
В качестве собственной задачи был составлен линейный график плотности генома Adhaeribacter radiodurans (Рисунок 8). Весь геном был поделен на участки, длиной 10 000 пар нуклеотидов каждый. С помощью методов и формул электронных таблиц Google Sheets (см. таблицу S1 сопроводительных материалов) было подсчитано, сколько генов содержится в каждом таком "кармане". Единственное важное замечание: гены, которые начинались в одном, а заканчивались в другом промежутке, считались в обоих из них. Самым густым оказался 174 (1 730 000 - 1 740 000 п.н.) - на нем было обнаружено 12 генов.
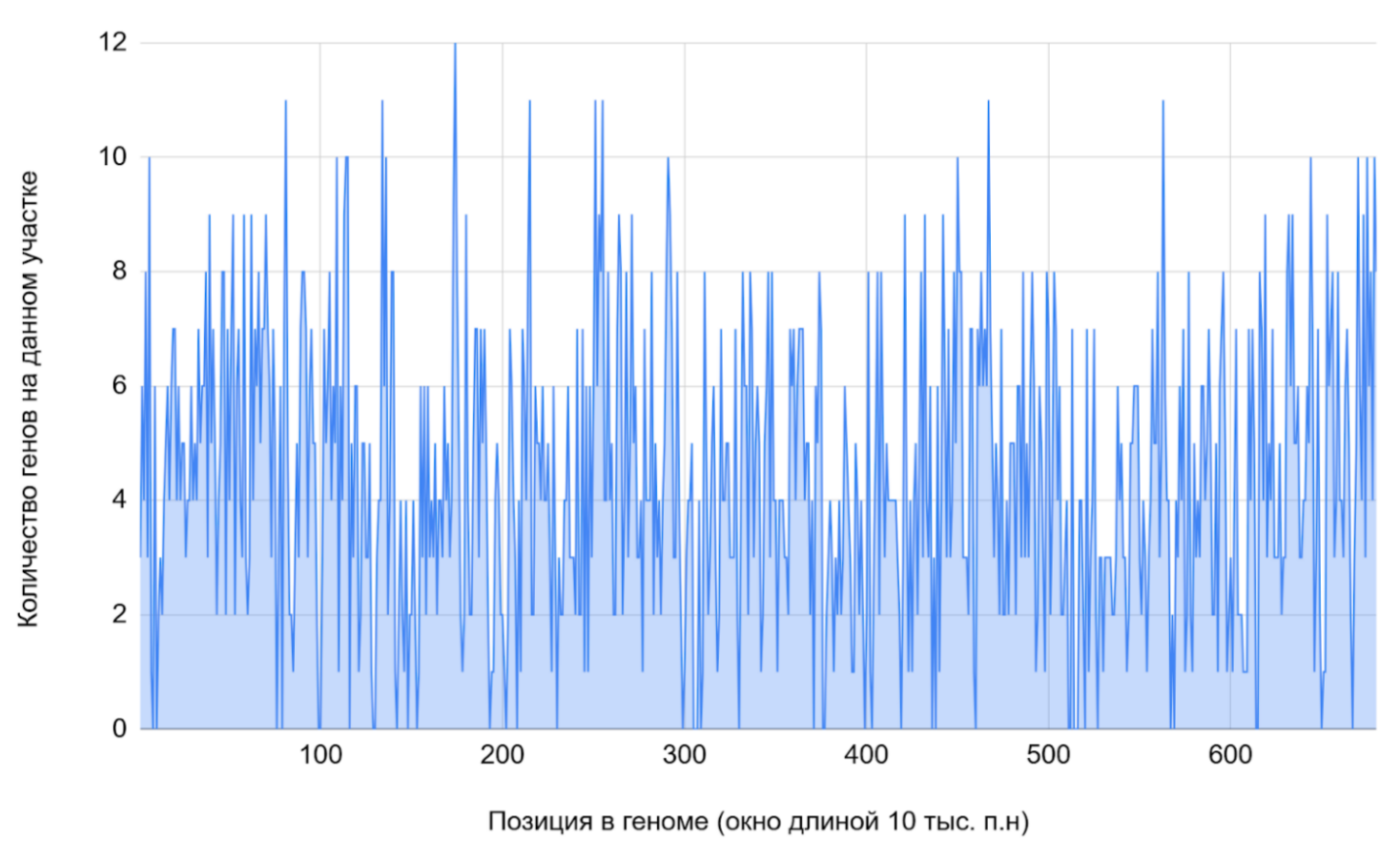
Также была составлена таблица отражающая количество участков низкой, средней и высокой плотности соответственно и вычислены некоторые статистические показатели для более детального анализа. Больше всего участков с низкой плотностью (тремя или четырьмя генами на промежутке) - их 101, а меньше всего - пустых: 22 участка (Таблица 6).
Таблица 6. Статистические показатели плотности генома
| Статистический показатель | Значение |
|---|---|
| Самый густой участок | 174 (на нем расположено 12 генов) |
| Медианная плотность | 4 |
| Средняя плотность | 4,47 |
Таблица 7. Сравнительная таблица плотности геномов прокариотов [13]
| Организм | % межгенной/регуляторной ДНК |
|---|---|
| Escherichia coli | 11-13% |
| Deinococcus radiodurans | 8-9% |
| Streptomyces coelicolor | 10-12% |
| Methanocaldococcus jannaschii | 7-9% |
1. Исходя из показателей плотности генома других прокариотов, (Таблица 7) можно сказать, что в геноме A. radiodurans довольно мало некодирующих участков (только 6.4% без генов)(Таблица 7). 2. Экстремальные контрасты — от 0 до 9+ генов в соседних окнах. 3. Широкое плато в середине — нет четкого стандарта распределения, следовательно, оптимальной плотности генома
Выводы
- Распределение длин белков типично для бактерий (медиана 288 а.о.), но присутствуют экстремально длинные белки (до 5441 а.о.), связанные с адгезией, формированием пилей и межклеточными взаимодействиями;
- Межгенные промежутки имеют медиану 250 п.н., что характерно для компактных геномов, выявлены значительные перекрывания генов (пик 0-5 п.н.);
- GC-состав (пик 43-44%) ниже, чем ожидается для радиорезистентных бактерий, что ставит под сомнение классическую радиорезистентность и указывает на адаптацию к ризосфере;
- Плотность генома неравномерна: 6.4% генома не содержат генов, выявлены сверхплотные кластеры (до 12 генов на 10 000 п.н.);
- Полученные данные свидетельствуют об адаптации бактерии к жизни в ризосфере, а не о специализации к радиорезистентности.
Сопроводительные материалы
Список литературы
- [1] Hwang YI, Son JS, Lee SY, Kim MJ, Park JM, Shin JH, Ghim SY. Adhaeribacter radiodurans sp. nov., isolated from the rhizospheric soil of Elymus tsukushiensis, a plant native to the Dokdo Islands, Republic of Korea. Int J Syst Evol Microbiol 2021; 71:4612.
- [2] LPSN Species: Adhaeribacter radiodurans
- [3] Yee, Mon Oo; Kim, Peter; Li, Yifan; Singh, Anup K.; Northen, Trent R.; Chakraborty, Romy. Specialized Plant Growth Chamber Designs to Study Complex Rhizosphere Interactions. Frontiers in Microbiology. 12 625752, 26 March 2021.
- [4] Таксономия бактерии была взята с сайта NCBI TAXONOMY BROWSER
- [5] Sergey Hosid, Edward N, Trifonov and Alexander Bolshoy. Sequence periodicity of Escherichia coli is concentrated in intergenic regions. BMC Molecular Biology 26 August 2004, 5:14
- [6] Owen M. Woodward and Terry Watnick Molecular. Structure of the PKD Protein Complex Finally Solved. Commentary on Su Q, Hu F, Ge X, et al. Structure of the human PKD1-PKD2 complex. Science. 2018;361(6406).
- [7] He Yang, Dong Wang, Li Guo, Huairong Pan, Robert Yvon, Scott Garman, Hen-Ming Wu, Alice Y. Malectin/Malectin-like domain-containing proteins: A repertoire of cell surface molecules with broad functional potential. CheungThe Cell Surface Volume 7, December 2021, 100056
- [8] Veggiani, Bijan Zakeri, Mark Howarth. Superglue from bacteria: unbreakable bridges for protein nanotechnology. Gianluca Trends in Biotechnology. Volume 32, Issue 10, October 2014, Pages 506-512
- [9] Georg Beckmann, Jens Hanke, Peer Bork, Jens G. Reich. Merging extracellular domains: fold prediction for laminin G-like and amino-terminal thrombospondin-like modules based on homology to pentraxins. Journal of Molecular Biology, Volume 275, Issue 5, 6 February 1998, Pages 725-730
- [10] Lijuan Gao Yahong Tan Weican Zhang Qingsheng Qi Xuemei Lu. Cytophaga hutchinsonii SprA and SprT Are Essential Components of the Type IX Secretion System Required for Ca2+ Acquisition, Cellulose Degradation, and Cell Motility. Front. Microbiol., 12 February 2021
- [11] Galtier N, Lobry J.R. Relationships between genomic G+C content, RNA secondary structures, and optimal growth temperature in Prokaryotes. Journal of Molecular Evolution. 44 (6): 632-636.
- [12] John P. McCutcheon, Bradon R. McDonald, Nancy A. Moran. Origin of an Alternative Genetic Code in the Extremely Small and GC-Rich Genome of a Bacterial Symbiont. Plos Genetics
- [13] Mira, A., Ochman, H. & Moran, N.A. Deletional bias and the evolution of bacterial genomes. Trends Genet. 17, 589-596 November 2001