В данном задании я работала со структурами 5ZC6, полученной методом ЯМР, и 121P, полученной методом РСА.
5ZC6 это структура раствора мутантного белка H-RasT35S в комплексе с KBFM123. Числопосчитанных моделей = 100, засабмиченных = 15.
121P - "структура и механизм гуаносинтрифосфатного гидролиза С-концевого укороченного белка рака человека P21-H-RAS (язык названия белка на странице PDB
определен как люксембургский, поэтому перевод с него на английский и потом на русский). Разрешение 1.54 Å.
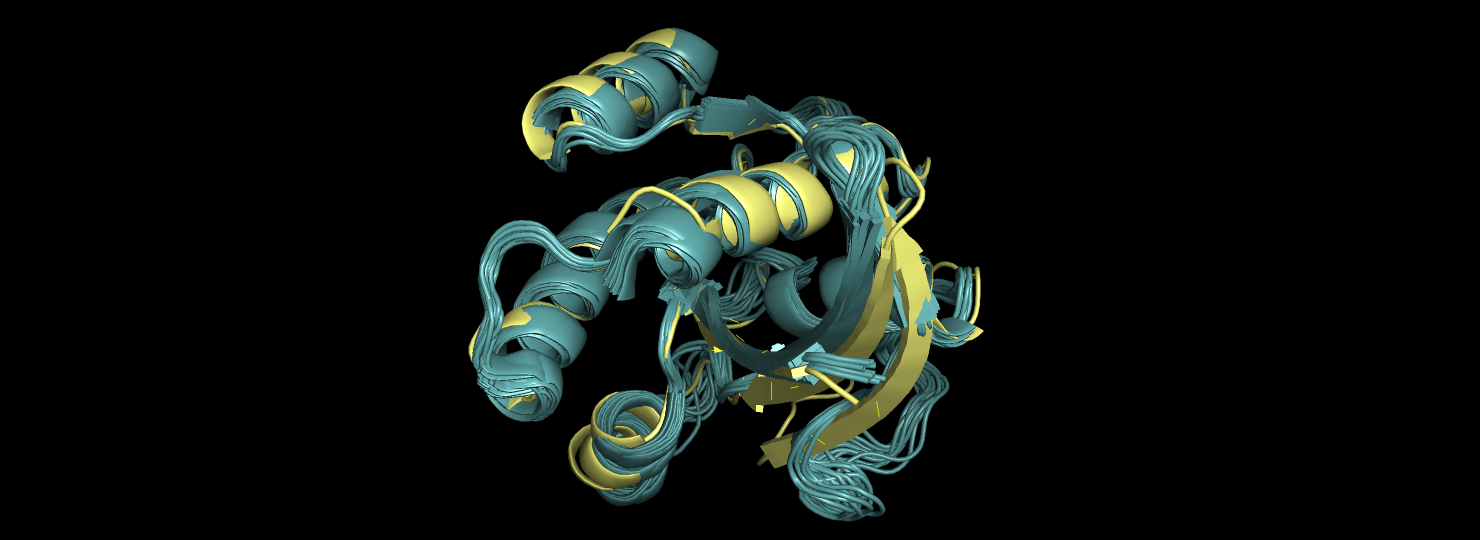
Рассмотрим выравнивания двух структур (1 РСА и 15 моделей ЯМР), представленных на рис. 1-4.
Видно, что в целом оба метода описывают не разные белки, но в большинстве неупорядоченных регионах и в некоторых бета-структруах, которые находятся на
поверхности молекулы, положение остатков различаются (рис. 1-2). Это связано с подвижностью остатков в этих областях.
В таких областях с низкой сходимостью моделей мы мало что можем сказать о положении атомов.
Рис.1.Выравнивание 5ZC6 (серо-голубой) и 121P (желтый).
Рис.2. Выравнивание 5ZC6 (серо-голубой) и 121P (желтый).
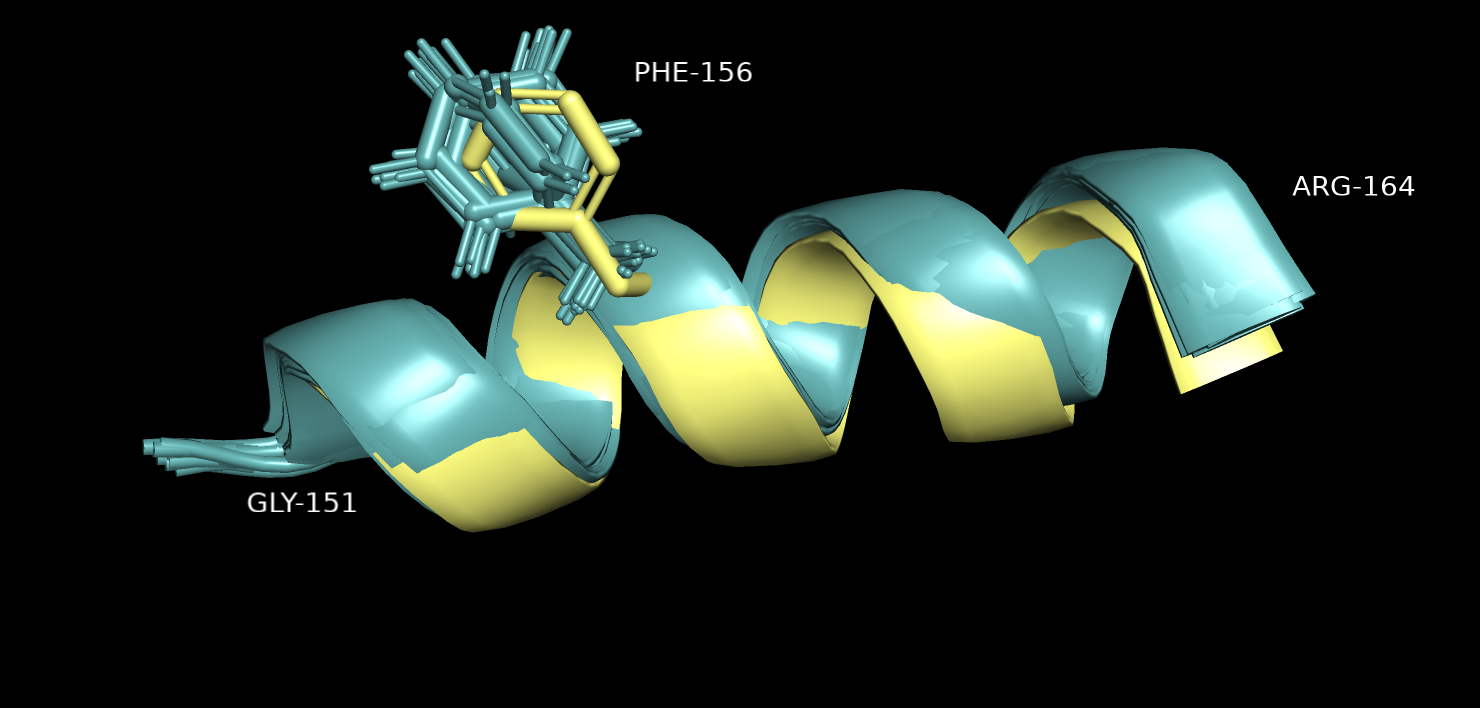
Рассмотрим различия на микроуровне. На рис. 3 и 4 показан участок одной альфа-спирали. Хотя общее положение цепи у двух структур
показано одинаковым, можно заметить различия в положениях боковых радикалов отдельных аминокислотных остатков. Например, на рис. 3 видно,
что у остатка Phe-156 ароматическое кольцо повернуто под разными углами для разных ЯМР моделей (в целом, есть 2 группы положений,
на которые поделись модели), но ни один из них не совпадает с тем положением, которое получено с помощью РСА.
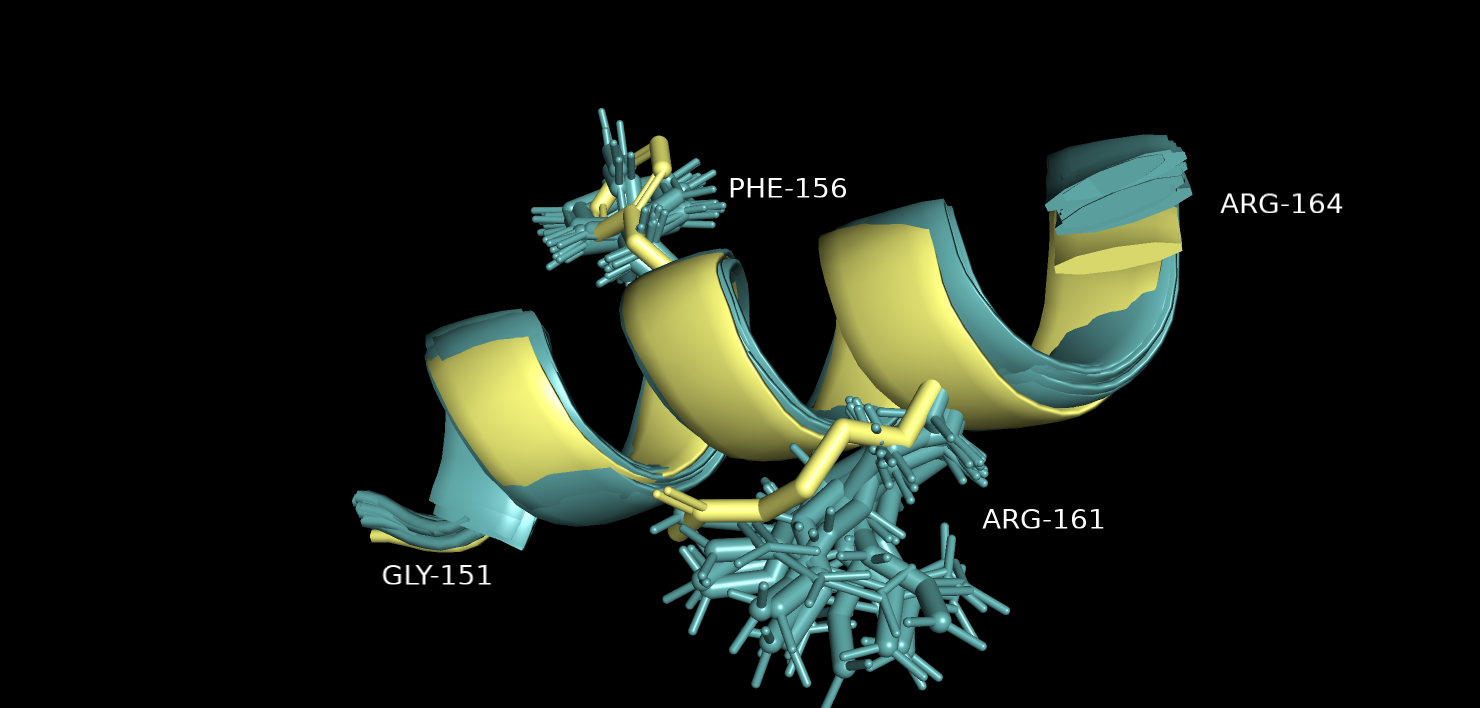
Также для остатка Arg-161 той же альфа спирали (рис.4), большинство моделей ЯМР для бокового радикала данного остатка не совпадают со структурой РСА.
Рис.3.Выравнивание 5ZC6 (серо-голубой) и 121P (желтый). Участок альфа-спирали
Рис.4. Выравнивание 5ZC6 (серо-голубой) и 121P (желтый). Участок альфа-спирали
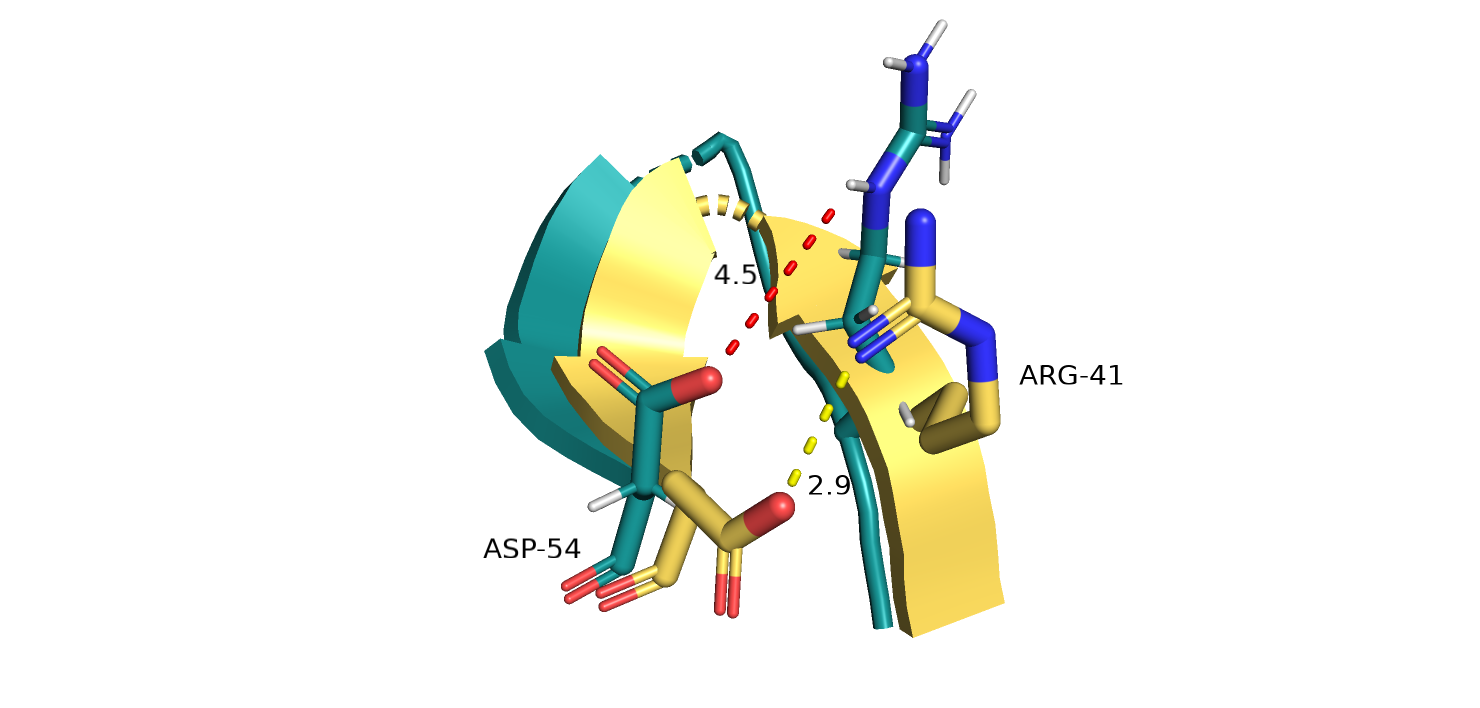
Другой пример различий на микроуровне. На рис. 5-6 показан участок бета-складчатости и 2 остатка на 2-х антипараллельных участках цепи.
На структуре, полученной методом РСА, можно предположить наличие водородной связи между остатками (показана желтым пунктиром на рис.6).
Для моделей ЯМР, в которых положение этих остатков сильно различается, в некоторых случаях возможно, но при усреднении моделей (рис. 6),
расстояние между остатками становится слишком большим для образования водородной связи. Возможно, данные остатки в белке довольно подвижны,
и водородная связь в данном случае не обязательно образуется, но так как сходимость моделей в данном случае не идеальное, однозначно утверждать
это нельзя.
Рис.5.Выравнивание 5ZC6 (серо-голубой) и 121P (желтый). Участок вета-структуры со всеми моделями ЯМР
Рис.6. Выравнивание 5ZC6 (серо-голубой) и 121P (желтый). Участок вета-структуры с одной моделью ЯМР.
Задание 2. RMSF.
Данное задание (и часть следующего) выполнено в файле notebook.
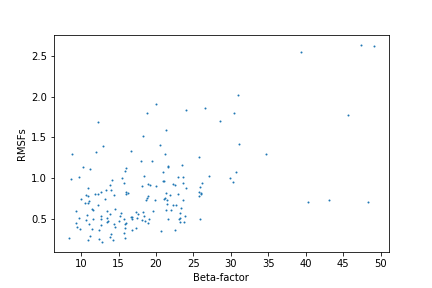
На рис. 7 мы можем наблюдать зависимость B-фактора аминокислотных остатков, полученных из РСА структуры, и RMSF из ЯМР структуры. В целом,
если не обращать внимание на аутлеты, можно представить линейную зависимость, так как чем меньше бета-фактор, тем меньше RSMF.
Большая часть точек располагается в области небольших значений обоих показателей. Это может быть связано как с небольшой подвижностью и
стабильностью белка, а также с тем, что при оптимизации моделей мы не можем нормально вписать в ЭП атом если он очень подвижен и выбрать очень подвижные
атомы, ухудшающие сходимость моделей. Таким образом, очень подвижные атомы мы не можем описать при методах ЯМР и РСА.
Рис.7. Зависимость RSMF от бета-факторов
Задание 3.
В данном задании мы считаем, что ансамбль моделей
отражает вариацию в позициях, которая появляется вследствие подвижности белка во времени. Я изучала водородные связи в 3 парах аминокислотных
остатков, представленных на следующих рисунках и в таблице.
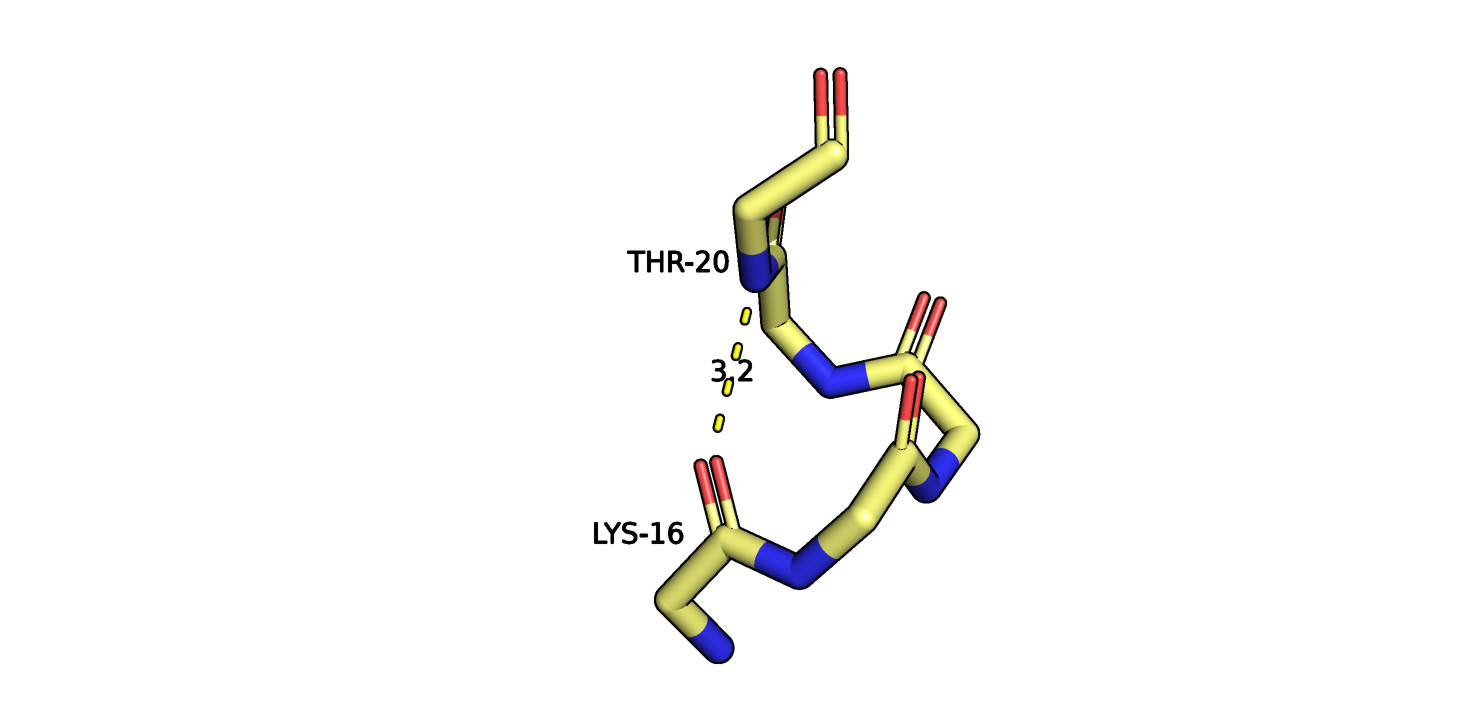
Рис.8. Остатки THR20-LYS16 121P (желтый) и их взаимодействие.
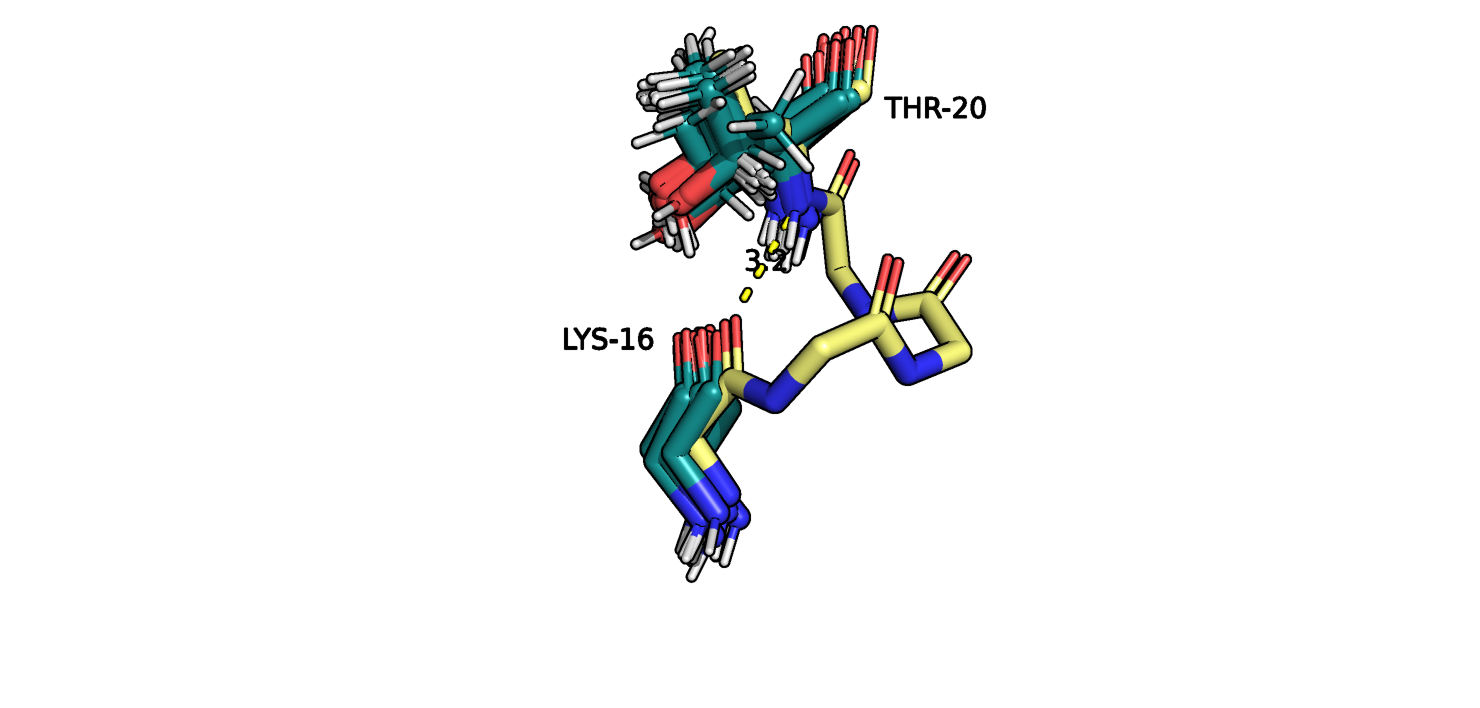
Рис.9. Выравнивание 5ZC6 (серо-голубой) и 121P (желтый). THR20-LYS16
На рисунках 8-9 изображен участок альфа спирали в ядре глобулы белка, пунктиром отмечено взаимодействие атомов остова остатков THR20 LYS16.
Атомы боковых радикалов не показаны. Видно, что положение атомов остова во всех моделях ЯМР отличаются незначительно. Об этом же говорит и
представленность водородной связи между этими остатками (см. табл.). Учитывая допущение относительно моделей ЯМР, можно сделать вывод о том,
что атомы остова малоподвижны (на данном участке данной альфа-спирали).
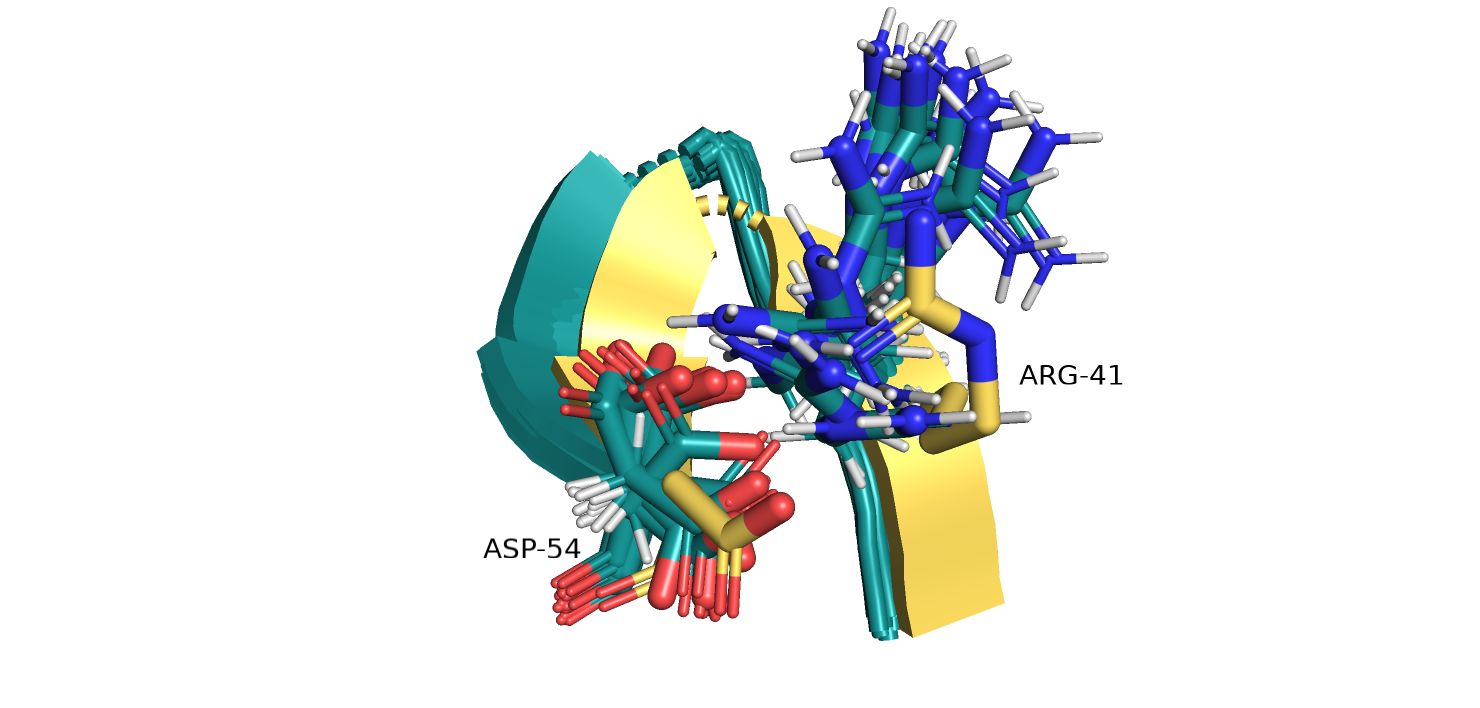
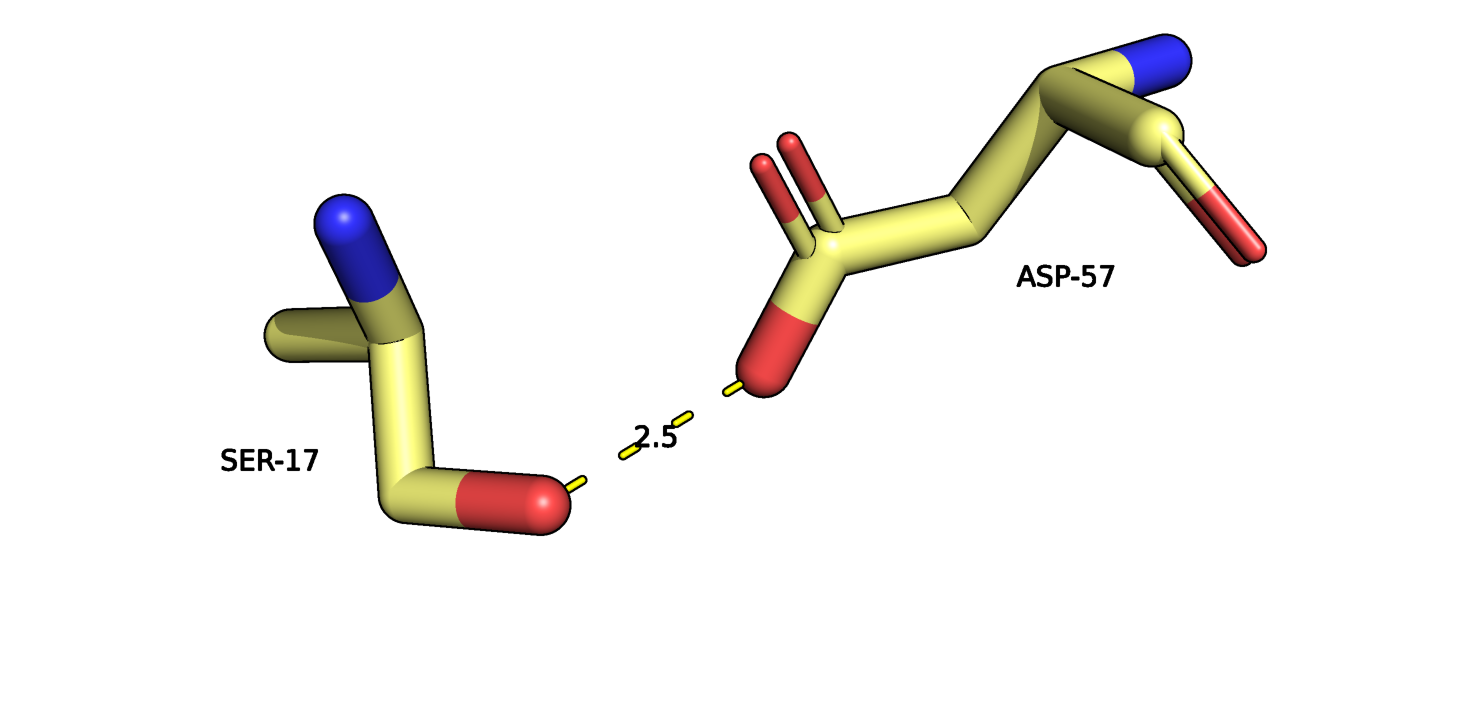
Рис.10. Остатки SER17 и ASP57 121P (желтый) и их взаимодействие.
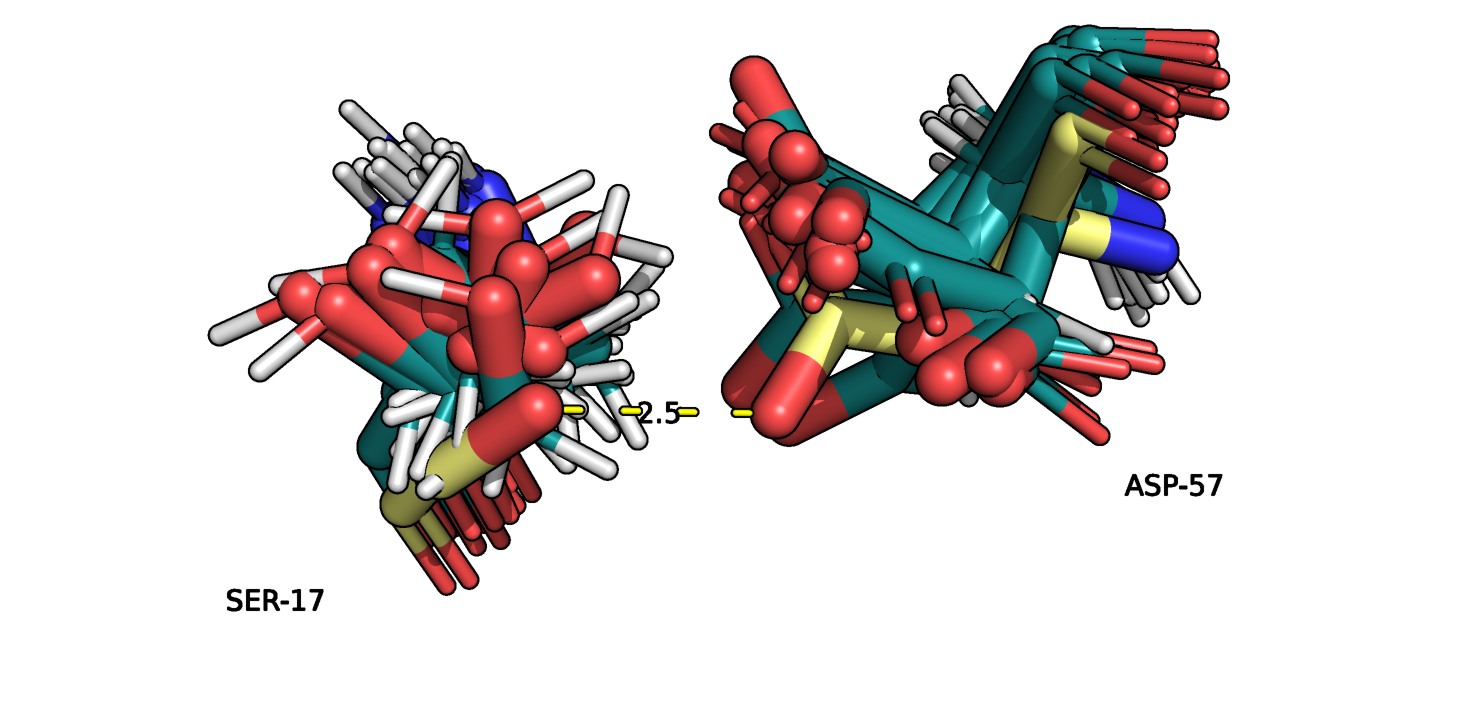
Рис.11. Выравнивание 5ZC6 (серо-голубой) и 121P (желтый). SER17 и ASP57
На рисунках 10-11 изображены SER17 и ASP57 из двух альфа спиралей в ядре глобулы белка,
пунктиром отмечено взаимодействие атомов боковых групп остатков. В данном случае вариантов расположения атомов для разных моделей заметно больше.
Одна и та же группа остатка может быть повернута в разные стороны, что говорит о ее большей подвижностью по сравнению с атомами остова.
Из 15 моделей только для 6 можно определить водородную связь в данном случае. При расчете количества водородных связей учитывался тот факт,
что при повороте на 180 градусов, атом O из OH-группы серина мог взаимодействовать как с O= группой так и с OH-группой аспарагина.
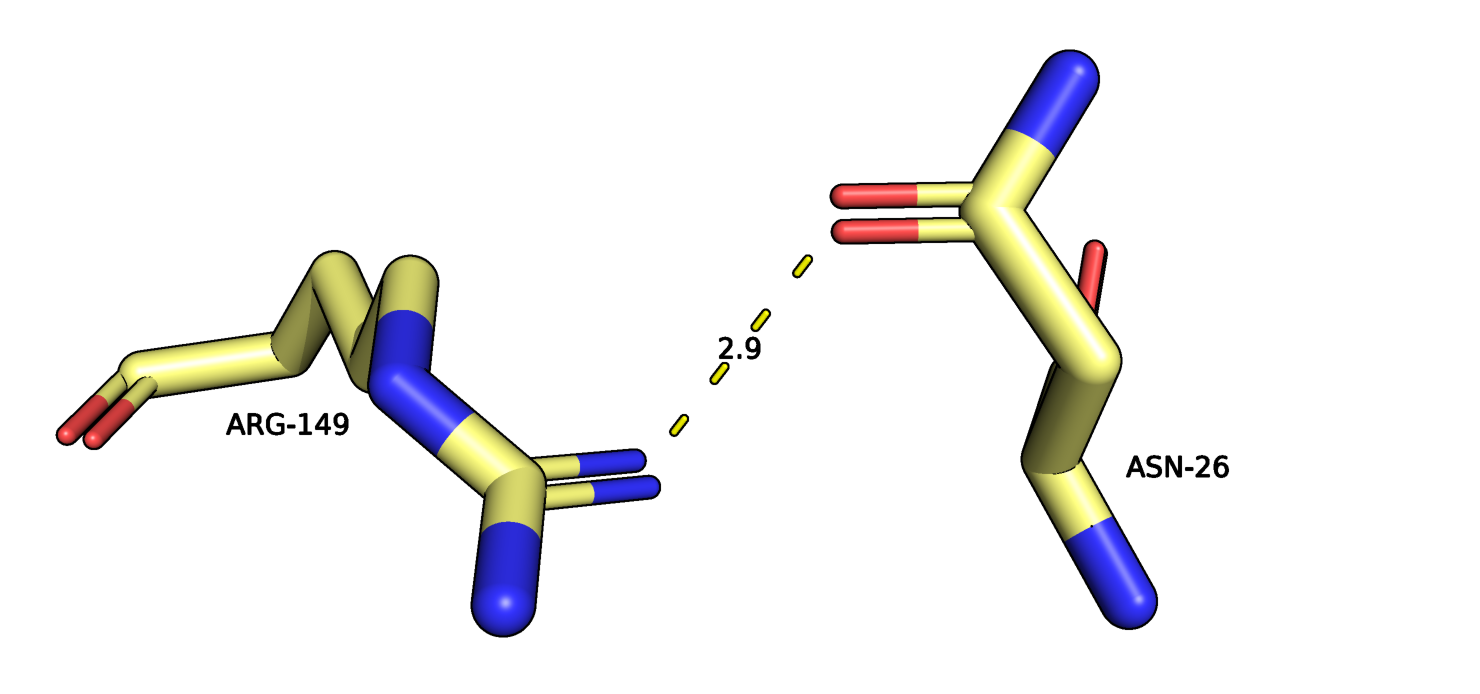
Рис.12. Остатки ASN26 и ARG149 121P (желтый) и их взаимодействие.
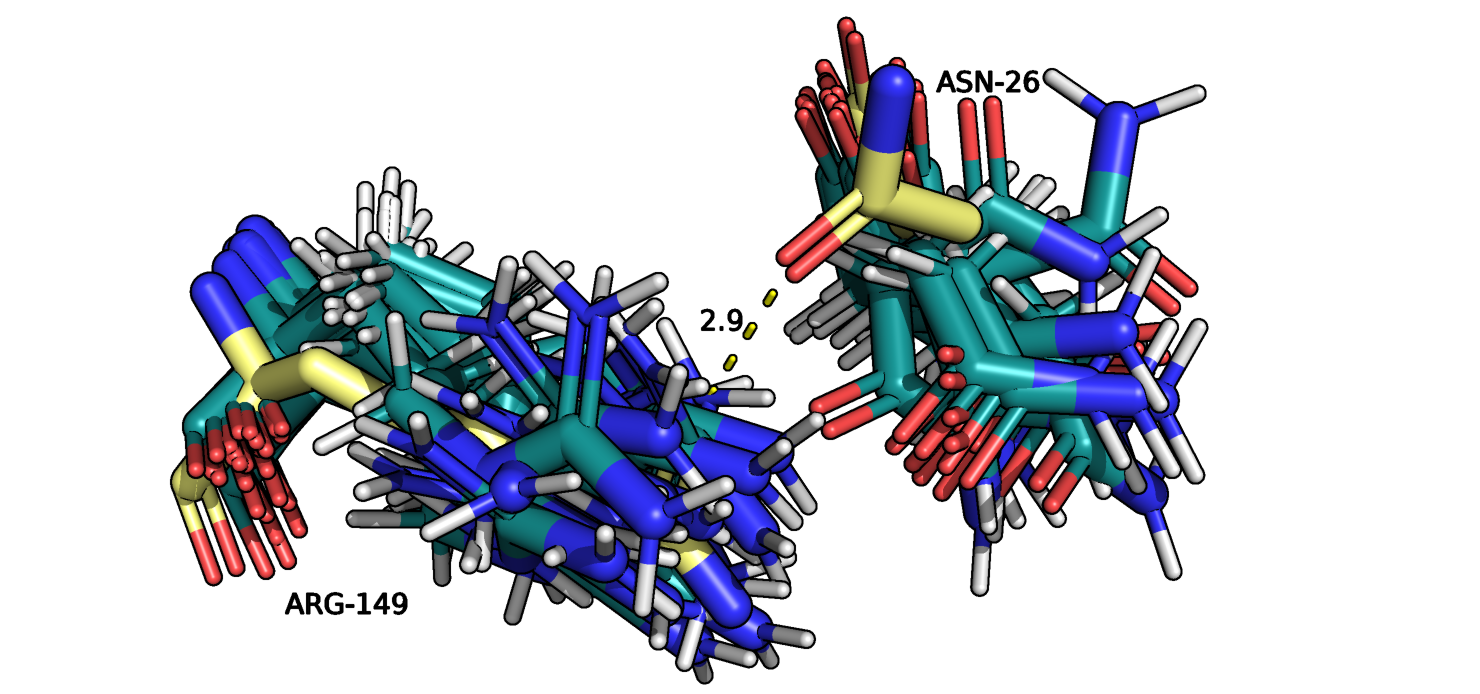
Рис.13. Выравнивание 5ZC6 (серо-голубой) и 121P (желтый). ASN26 и ARG149
На рисунках 12-13 изображены ASN26 и ARG149 из двух петлей на поверхности белка,
пунктиром отмечено взаимодействие атомов боковых групп остатков. Разнообразие расположения атомов для разных моделей сильно увеличилось.
Одна и та же группа остатка может быть повернута в разные стороны, что говорит о ее большей подвижностью по сравнению с атомами остова.
Из 15 моделей только для 4 можно определить водородную связь. При расчете количества водородных связей также учитывалась возможность
образования водородной связи с одним из двух атомов азота. В данном примере наблюдается самое большое значение (медиана и максимум) расстояния
между взаимодействующими (или не взаимодействующими по причини большого расстояния) атомами. Можно сделать вывод, что в неупорядоченных структурах (петлях),
положение которых слабо стабилизировано на поверхности белка, наблюдается наибольшая подвижность атомов.
При расчете количества водородных связей в ямр моделях, если расстояние между акцептором и донор было меньше 3.5 ангстрем, принималось, что
связь есть.
Пара остатков
Расстояние для РСА (121P)
Представленность связи в моделях ЯМР (5zc6)
min
max
med
Расположение
THR20-LYS16
3.2
12, 80%
3.21
3.64
3.36
ядро глобулы, остов альфа-спирали
SER17-ASP57
2.5
6, 40%
2.49
5.28
4.19
ядро глобулы, радикалы альфа-спиралей
ASN26-ARG149
2.9
4, 26.67%
2.58
5.80
4.09
на поверхности глобулы, остатки неупорядоченных петлей