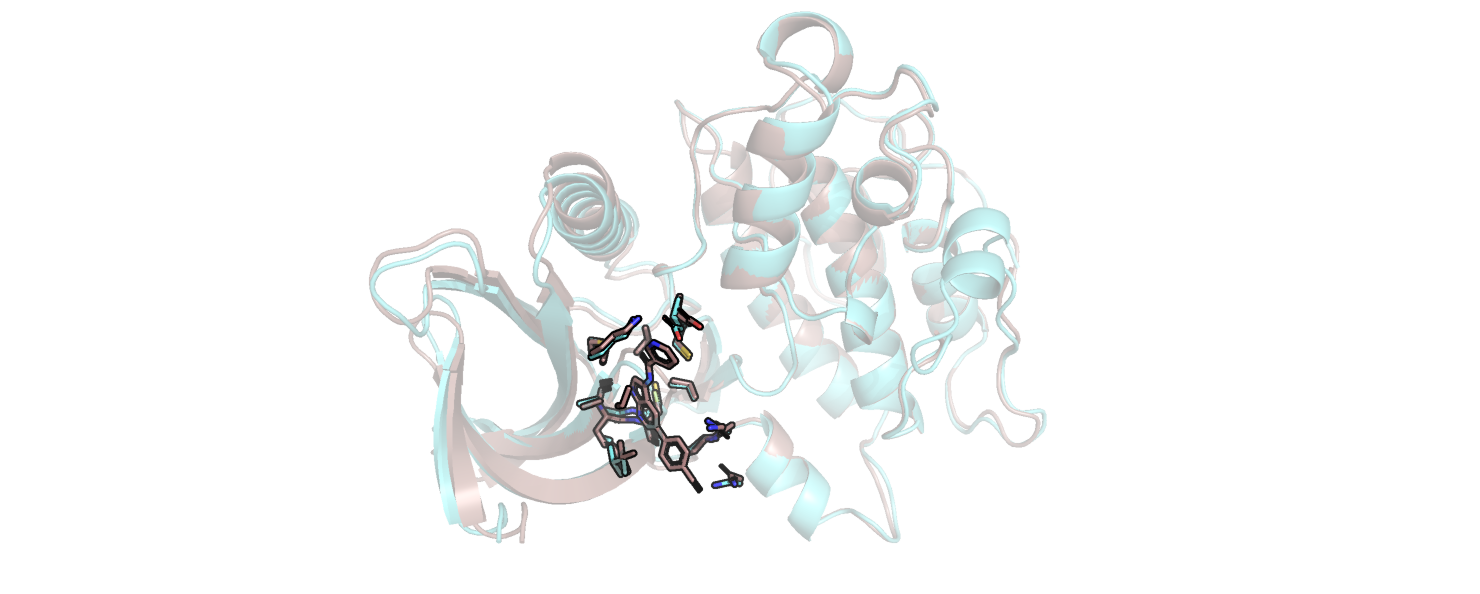
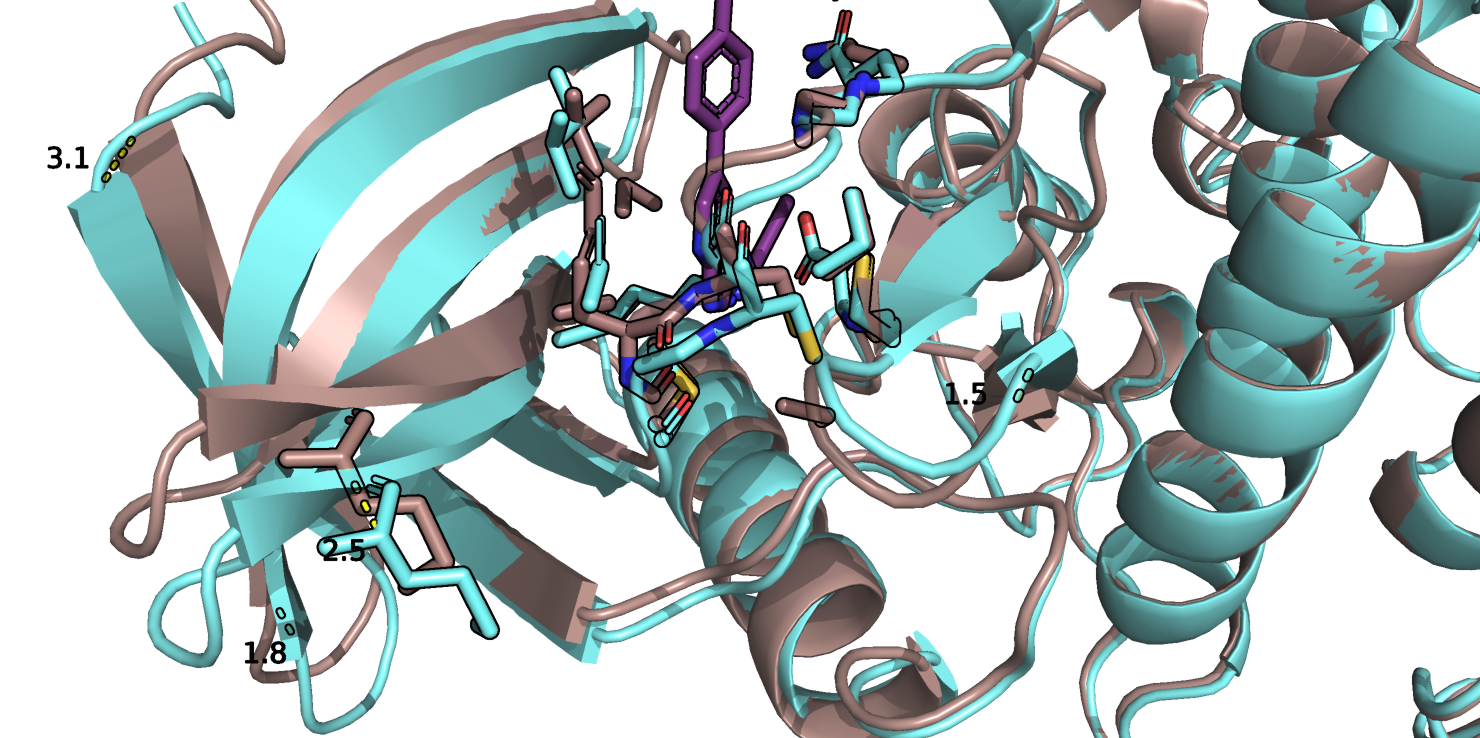В этом практикуме я изучала 2 структуры белка - связанную с лигандом и свободную. На рис. 1 голубым
покрашена свободная структура, коричневым - связанная.
Рис.1. Совмещенные структуры.
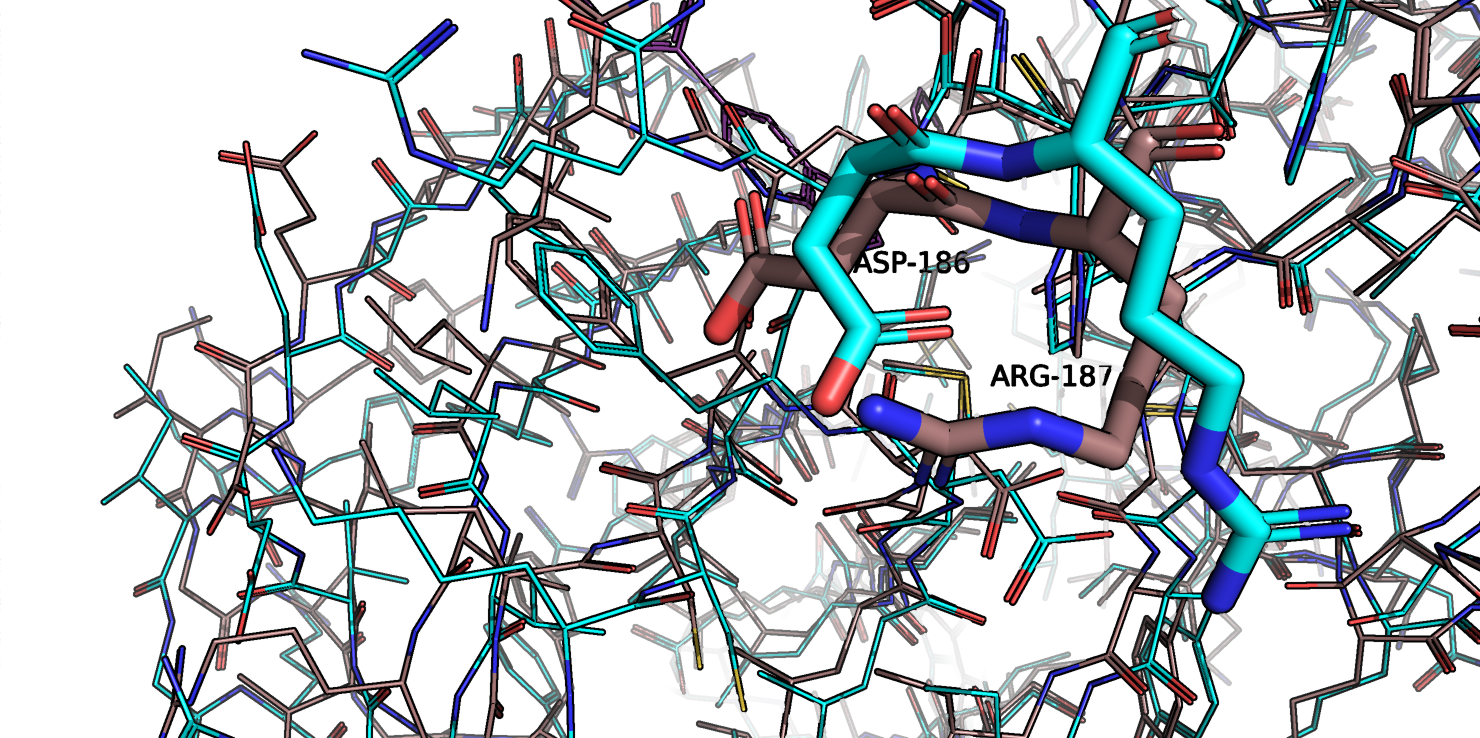
Рис.1b. Совмещенные структуры, фрагмент белка, приближенный к сайту связывания.
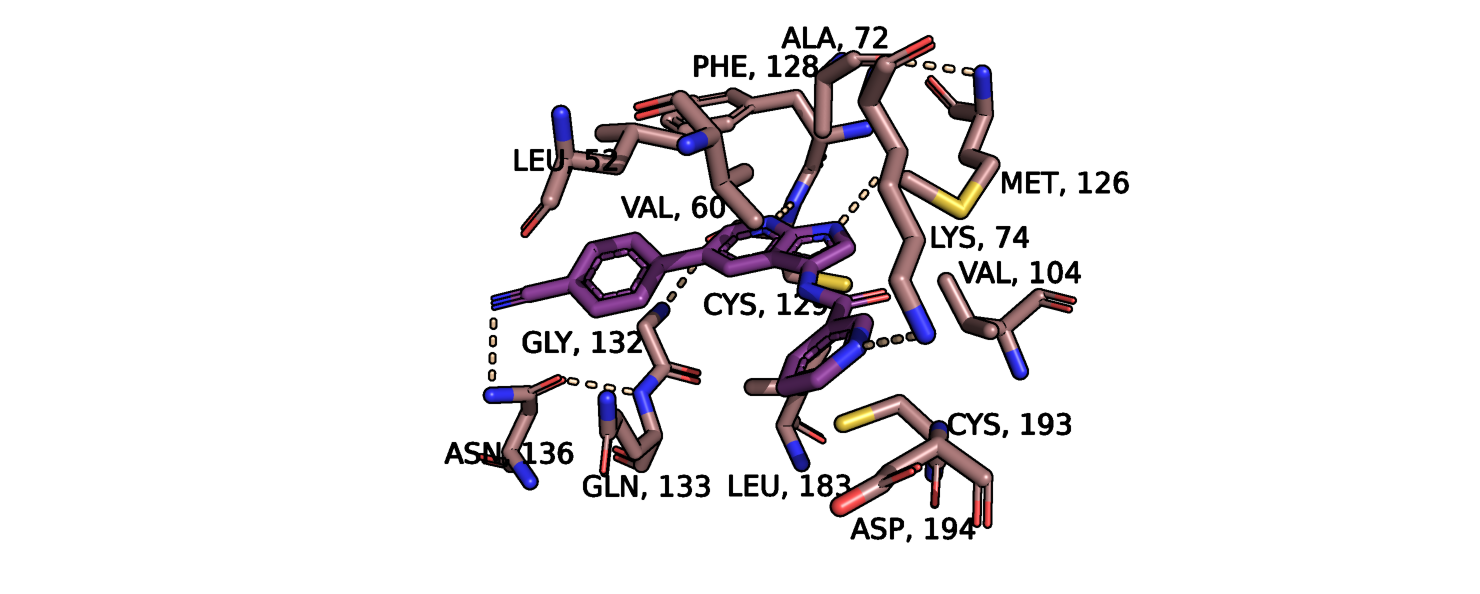
Рис.2. Совмещенные структуры. Сайт связывания с лигандом
Из рис.1 и 1b видно, что связывание лиганда немного меняет конформацию белка. В основном, немного смещается
та часть белка, которая ближе к сайту связывания - одна альфа спираль и бета-лист. В связанном состоянии альфа спираль и бета-тяжи немного
отодвигаются в сторону от лиганда на 1-2 ангстрем, а в самом отдаленном бета-тяже - на 3.1, хотя бета лист который находится рядом с лигандом
ближе в альфа-спиральному домену, сдвигается в ту же сторону что и другие бета-тяжи - т.е. подвигается ближе к лиганду. На уровне боковых радикалов аминокислот в сайте связывания лиганда также происходят изменения (рис.2). Сильнее всего изменяется
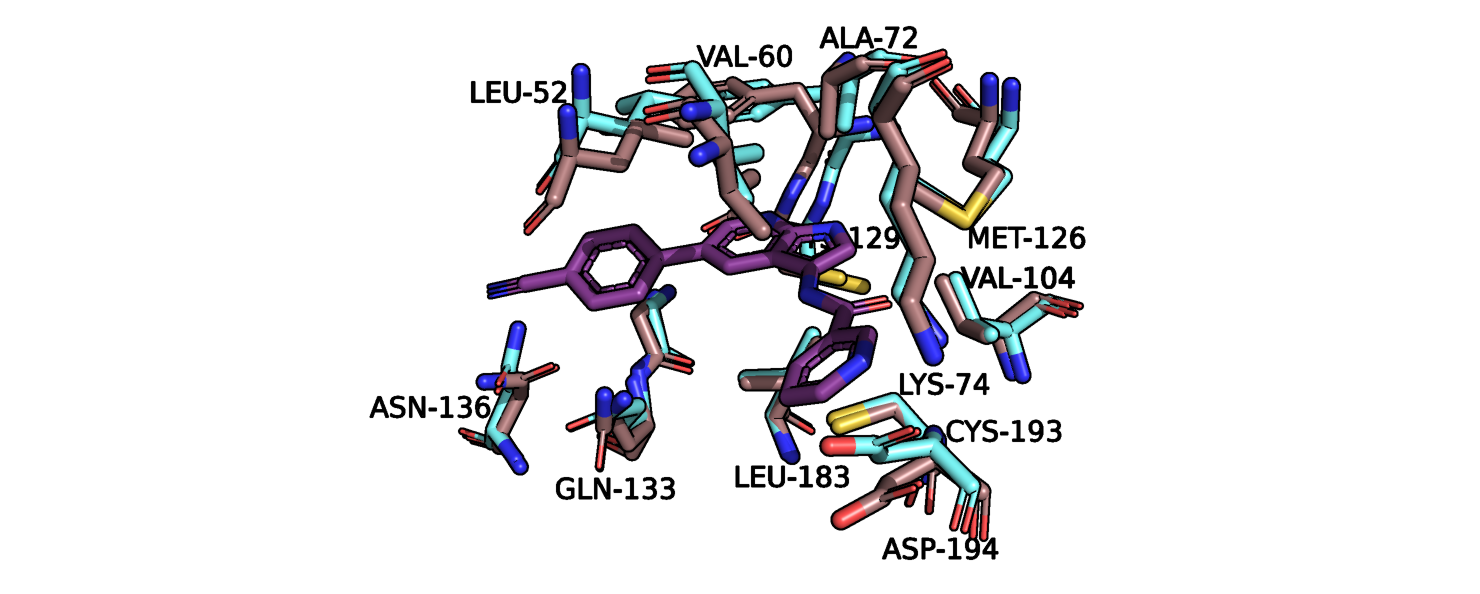
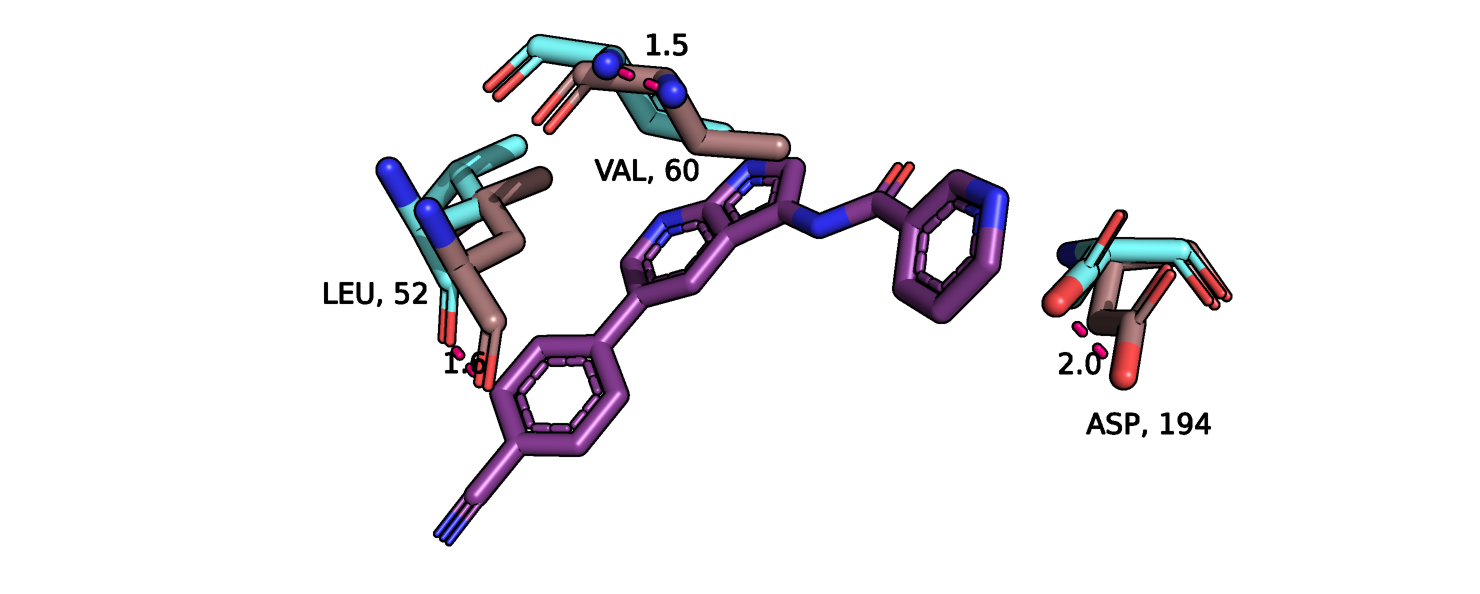
положение аспартата-194, валина-60, лейцина-52. Более детально эти остатки показаны на рис.5. Максимальное отклонение аспартата-194
составляет 2 ангстрема, причем в свободной конформации он отклоняется в сторону центра кармана связывания, а в присутствии лиганда - в обратную
сторону, увеличивая радиус кармана. Примечательно, что данный остаток и 2 других не образуют водородных связей с молекулой лиганда,
т.е. изменение его положения может влиять на способность лиганда покидать карман с помощью диффузии. Кроме того, два других остатка смещаются
в ту же сторону, что и данный аспарагин. Аспартат в свободной конформации может также препятствовать проникновению лиганда внутрь сайта связывания.
Изменения в конформации белка, по-видимому, связаны именно с появлением в кармане лиганда.
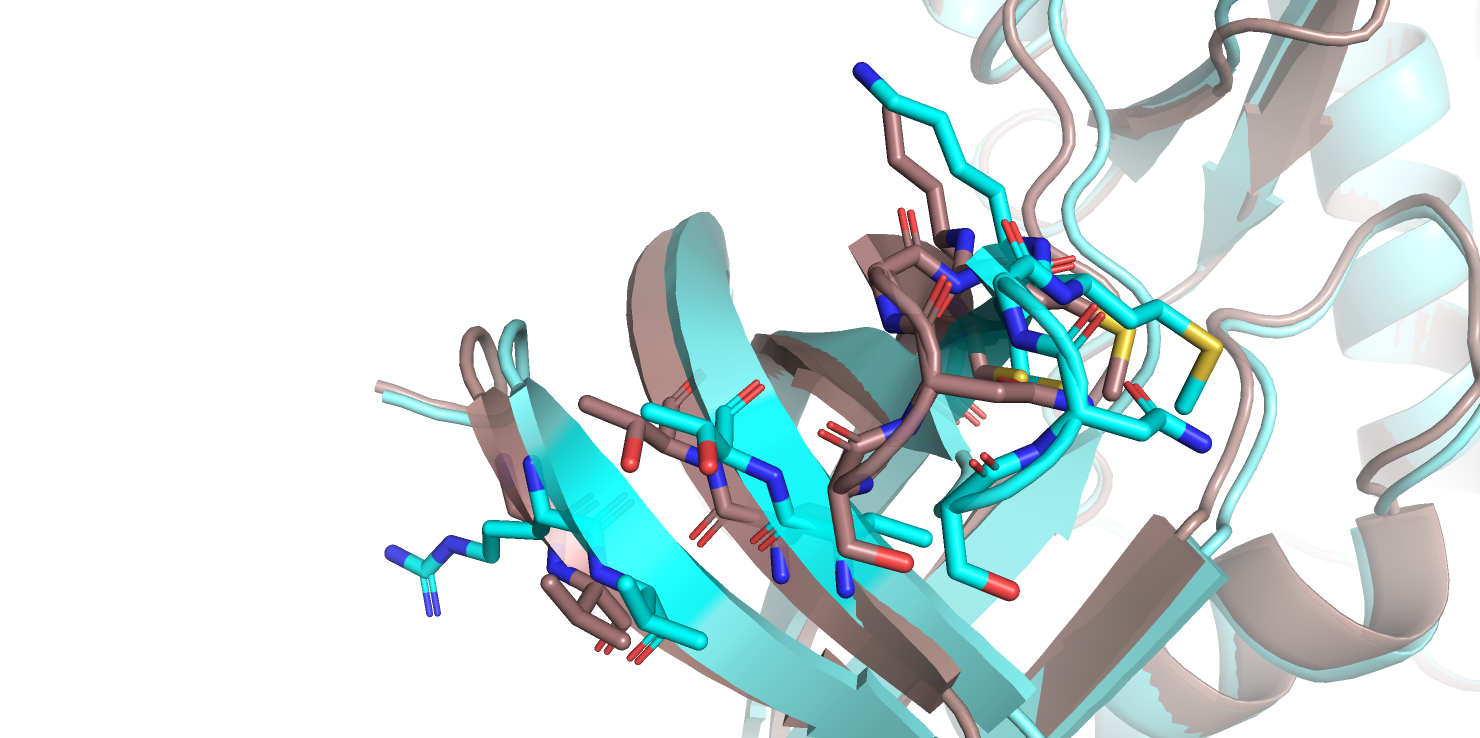
Если посмотреть на остатки во вторичных структурах (рис.1с), которые сильнее всего сместились, можно заметить, что они буквально просто сместились все в одну
сторону и каких-то новых взаимодействий я не смогла найти. А если у остатков и меняются углы, то новых взаимолействий все равно нет, так как такие остатки
оказываются на поверхности белка (рис. 1d).
Рис.1с. Совмещенные структуры. Смещение остатков в бета-листе
Рис.1b. Совмещенные структуры, изменение конформации остатков.
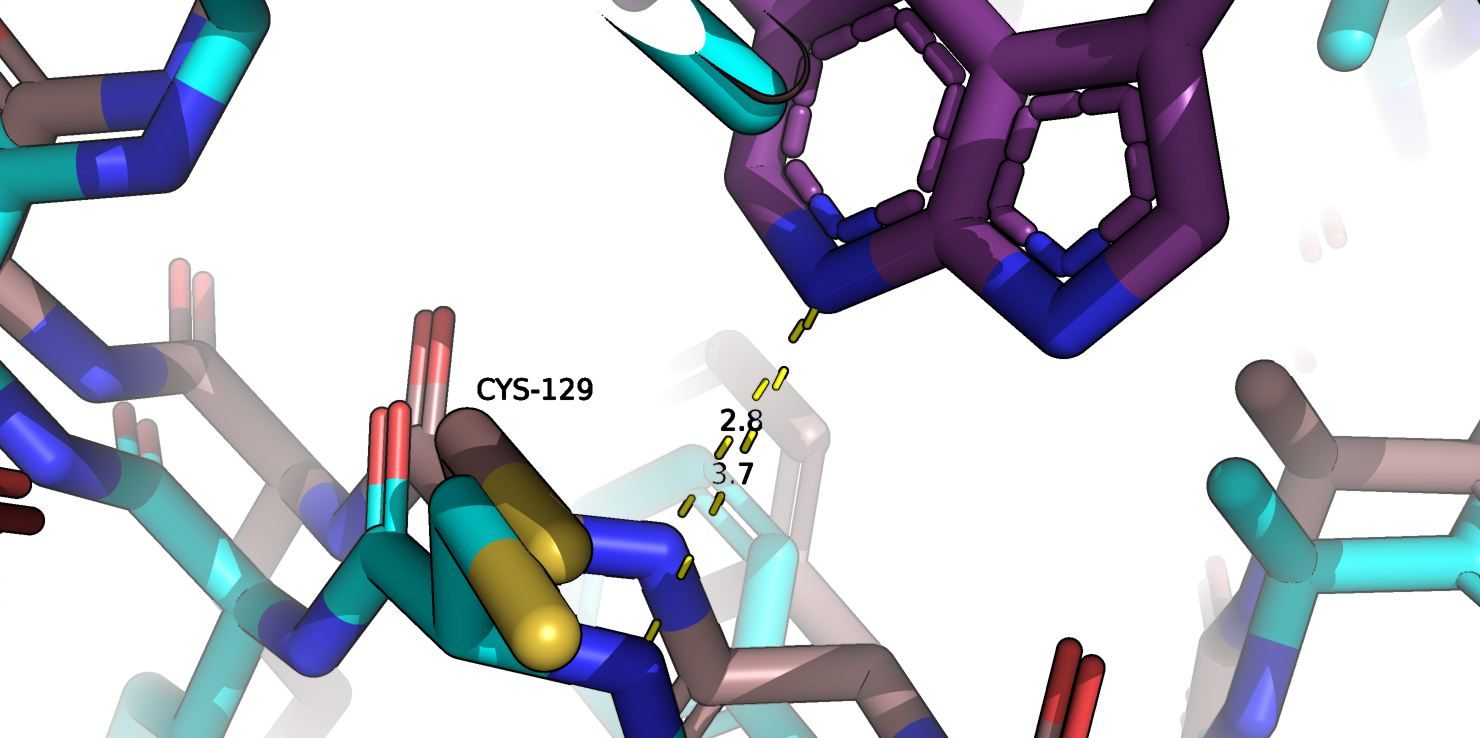
В самом сайте связывание изменение конформации в некоторых случаях можно обьяснить появлением связей с лигандом.
Например, на рис. 1e показано расстояние между взаимодействующими атомами лиганда и остатка цистеина-129. В свободной форме этот остаток отдаляется от кармана на прмиерно 0.9 А.
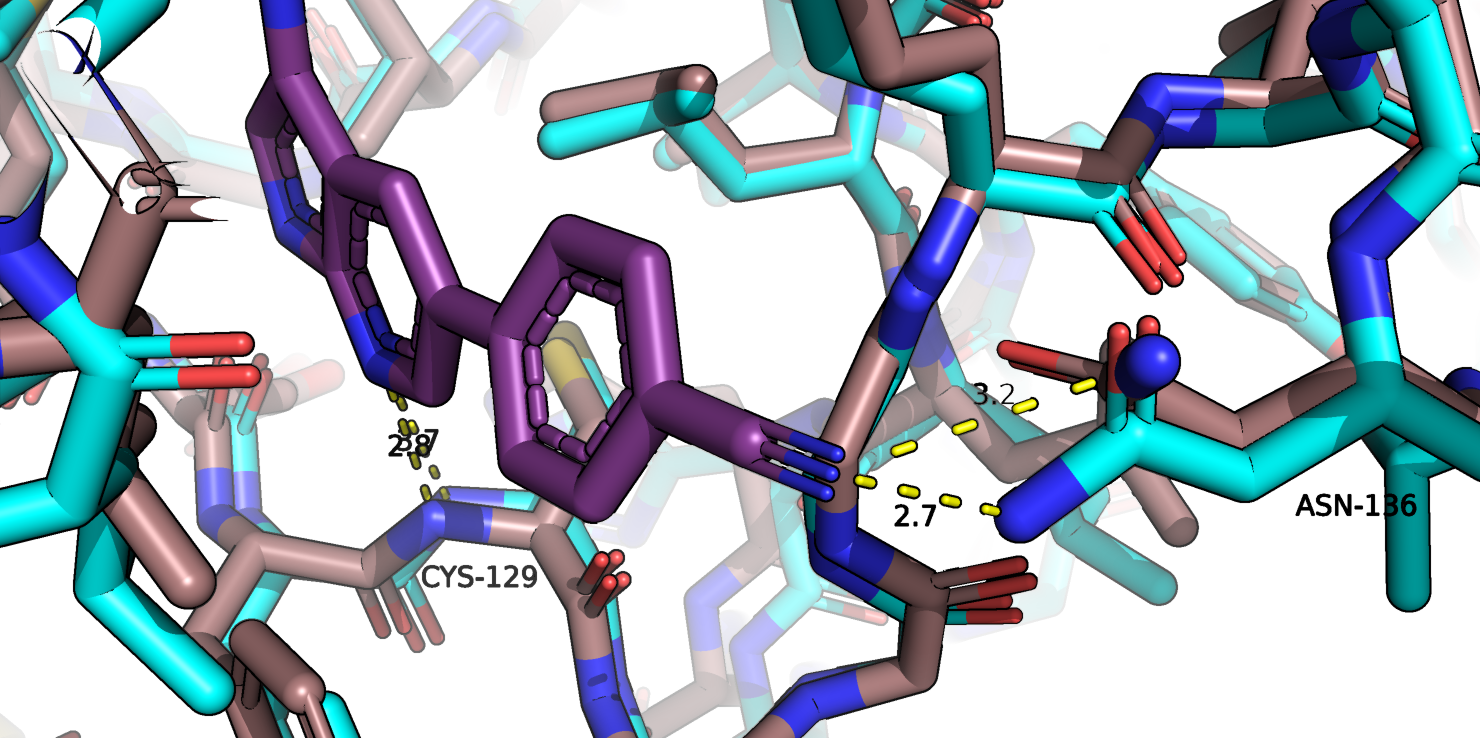
Но такое взаимодейсвтие, возможно, не совсем обьясняет изменение конформации на 3 А в других участках белка. Также на рис. 1f показано
взаимодействие с другим атомом лиганда, однако оно возникает в основном из-за поворота функциональной группы аспарагина-136 и тоже не обьясняет
смещение белка.
Рис.1с. Совмещенные структуры. Взаимолействие в кармане с лигандом цистеина-129
Рис.1b. Совмещенные структуры, взаимодействие с лигандом аспарагина-136.
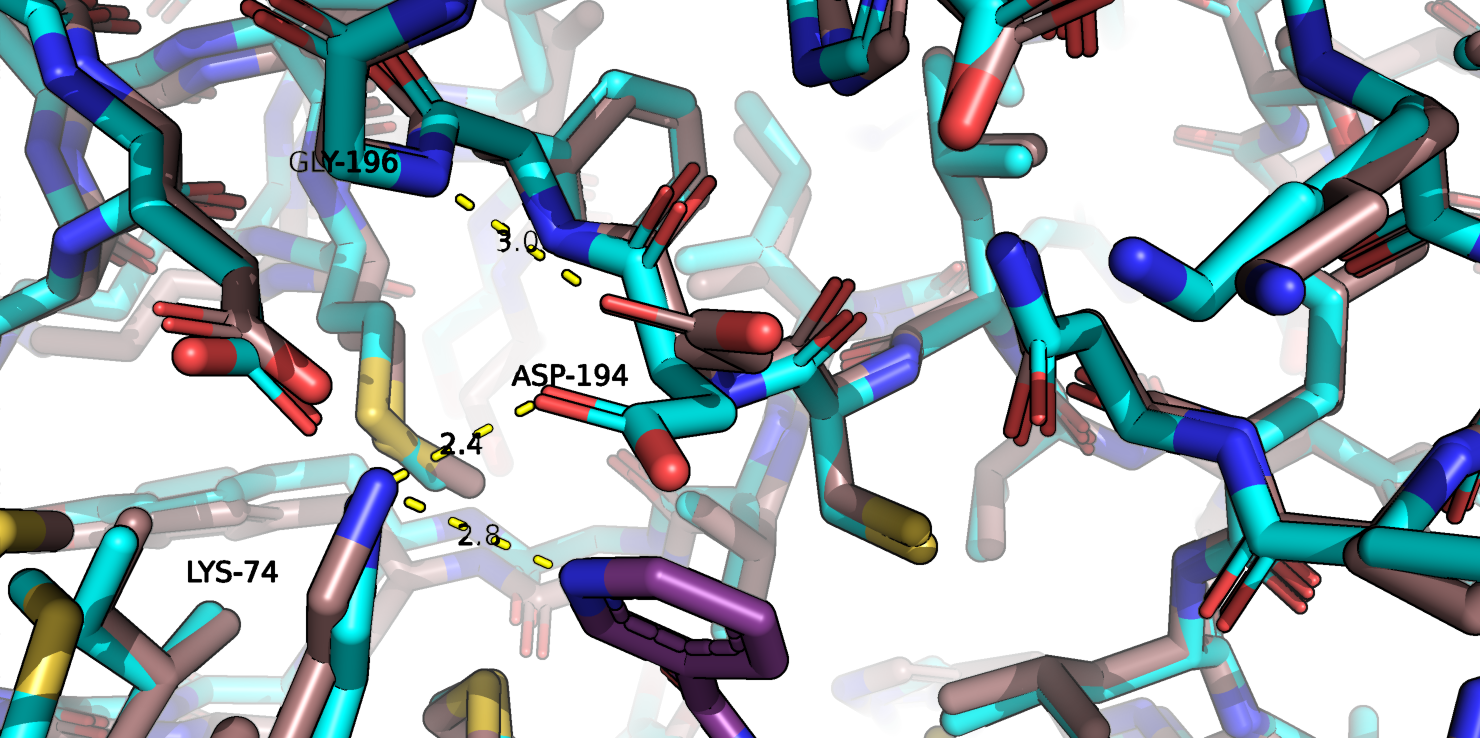
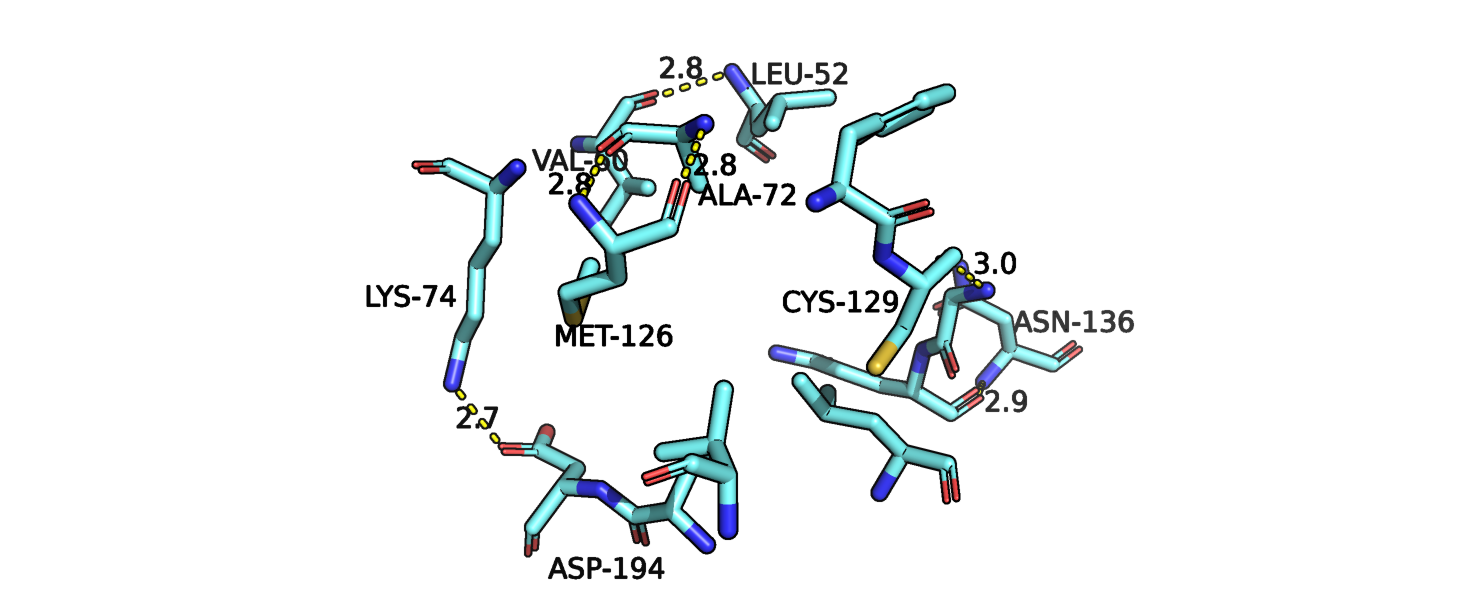
Вернемся к аспартату-194, о котором говорилось выше. Как уже говорилось, он не образует водородных связей с
лигандом и отклоняется от центра кармана при связывании лиганда. Исходя из того, что можно увидеть на рис.1j, можно предположить, что
в свободной форме остаток аспартата-194 может спокойно расположиться в кармане и, возможно, взаимодействовать с остатком лизина, однако
при связывании лиганда, аспартат отклоняется подальше, где может взаимодействовать с остатком глицина-196, который, кстати не меняет конформации,
а лизин теперь может взаимодействовать с лигандом и тоже практически не меняя свое положение.
Рис.1j. Совмещенные структуры. Взаимолействия в кармене связывания
Рис.3. Сайт связвания лиганда белка состоянии связанного лиганда.
Рис.4. Сайт связвания лиганда белка в свободном состоянии.
Рис.5. Сайт связвания лиганда, показаны остатки с наибольшим изменением конформации.
Из файла pdb со связанным лигандом я удалила строки, связанные с лигандом, загрузила этот файл в сервис POCASA и получила файл.
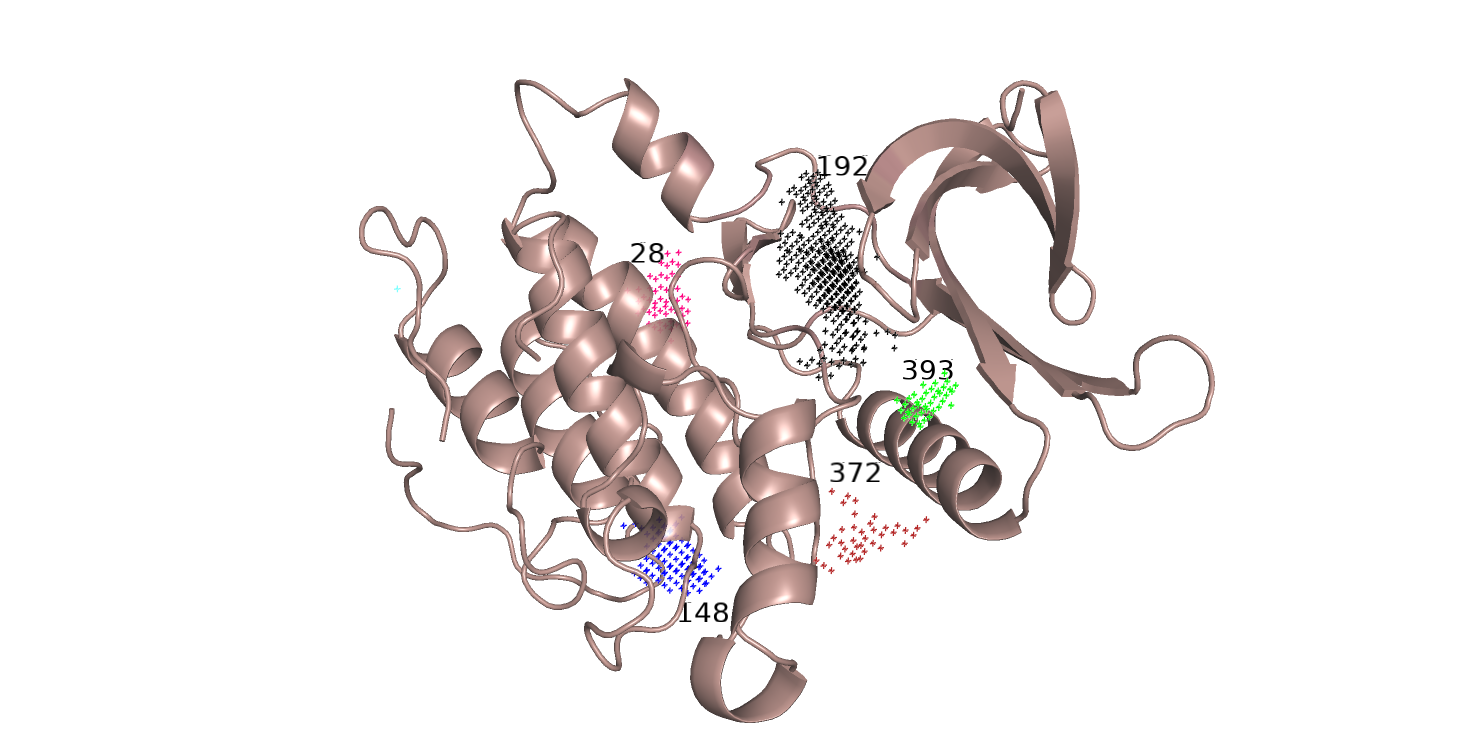
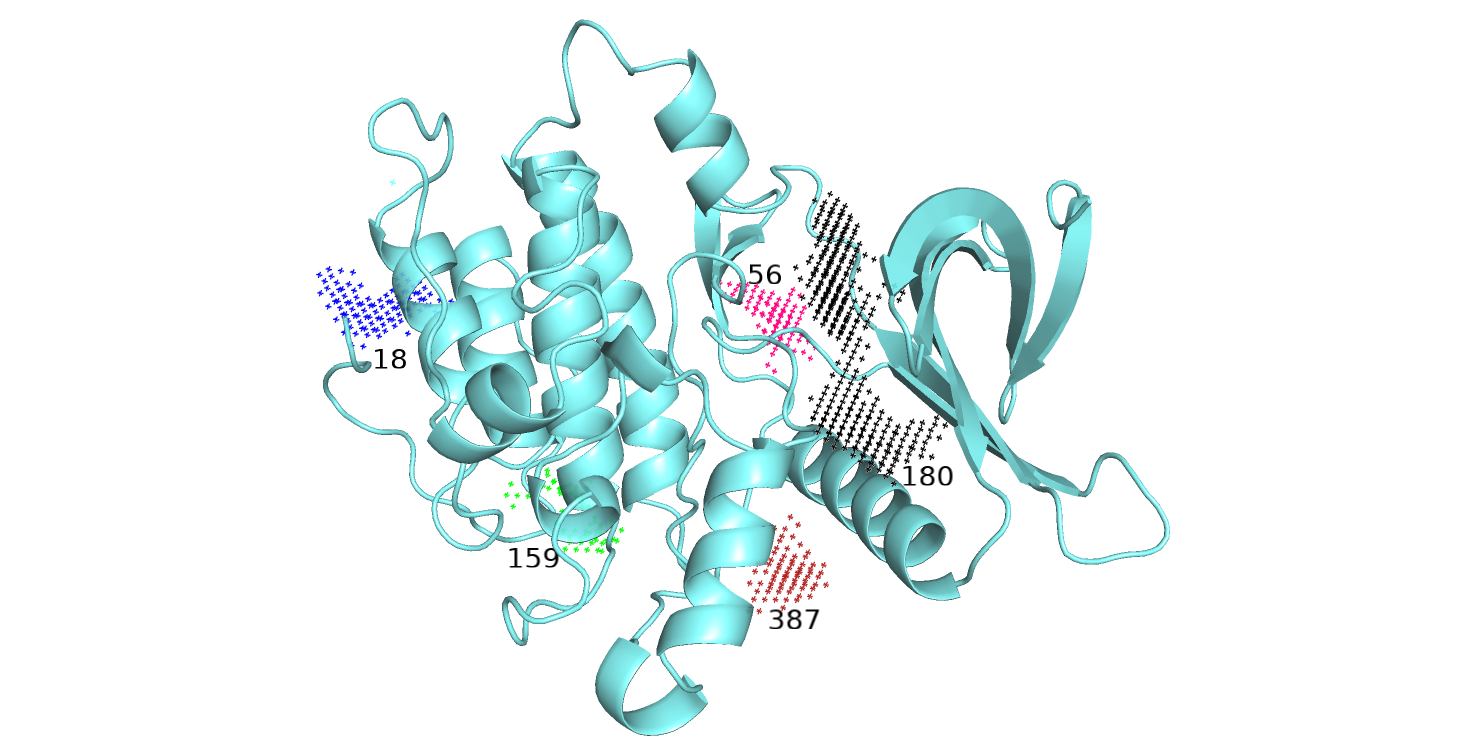
Топ-5 полученных карманов показаны на рис. 6 разными цветами. То же я проделала со "свободной" структурой.
Топ-5 ID карманов для этой структуры: 180, 18, 159, 56, 387.
Полость, в которой находится лиганд соответствует карману 192,т.е. первому в топе. Остальные карманы, как мне кажется, тоже
хорошо соответсвуют полостям на поверхности белка. Сравнивая карманы на рис. 6 и 7, можно заметить,
что 1-ый в топе карманов (черный) частично совпадает у белка, связавшего лиганд, и у белка без лиганда.
У структуры на рис. 6 этот карман кажется меньше, чем на рис. 7, однако можно предположить, что при изменении
конформации карман с ID 393 (зеленый) расширяется и становится частью кармана-180.
Возможно, это связано с тем, что в закрытой конформации с лигандом карман сужается для того, чтобы сильнее связать лиганд,м
а затем карман становится больше, чтобы повысить вероятность того, что лиганд попадет в этот карман.
Можно также предположить, что карман 372 и 387 это один и тот же карман, но из-за конформационных изменений,
при связывании лиганда этот карман немного расширяется. Синие и розовые карманы не совпадают между собой по расположению и, возможно,
соответсвуют нефункциональнозначимым полостям на поверхности белка. Также карман-159, возможно, образуется из 148-ого при
изменении конформации.
что карман
N Топ
ID
Объем
VD value
1
180
380
1234
2
18
99
264
3
159
60
255
4
56
92
218
5
387
74
186
Параметры карманов для "свободной" структуры
N Топ
ID
Объем
VD value
1
192
309
1061
2
148
88
281
3
393
40
145
4
28
55
137
5
372
39
107
Параметры карманов для "связанной" структуры
В представленных выше таблицах показаны параметры карманов для двух структур. Обьем
самой большой полости, в которой связывается лиганд, у белка с лигандом равен 309, а без лиганда - 380. Даже если
учитывать обьем кармана-393 (объем 40), как часть кармана-180, то объем кармана без лиганда получается больше. Это можно
обьяснить "стягиванием" лигандом остатков белка. Можно предположить, что когда лиганд попадает в карман,
он сначала стягивает остатки в сайте связывания, а это приводит к конформационному изменению остального кармана и всего белка.
Задание 2. Протонирование, подготовка к докингу
Для выполнения докинга сначала нужно добавить водороды в структуру белка в свободной конформации и в структуру лиганда, взятого из pdb файла
белка, связанного с лигандом. Протонированная структура белка была получена с помощью сервиса PDB2PQR. Лиганд я протонировала через SPORES
(рис. 8-9).
Рис.8. Лиганд без водородов
Рис.9. Лиганд с добавленными водородами
Задание 3. Докинг
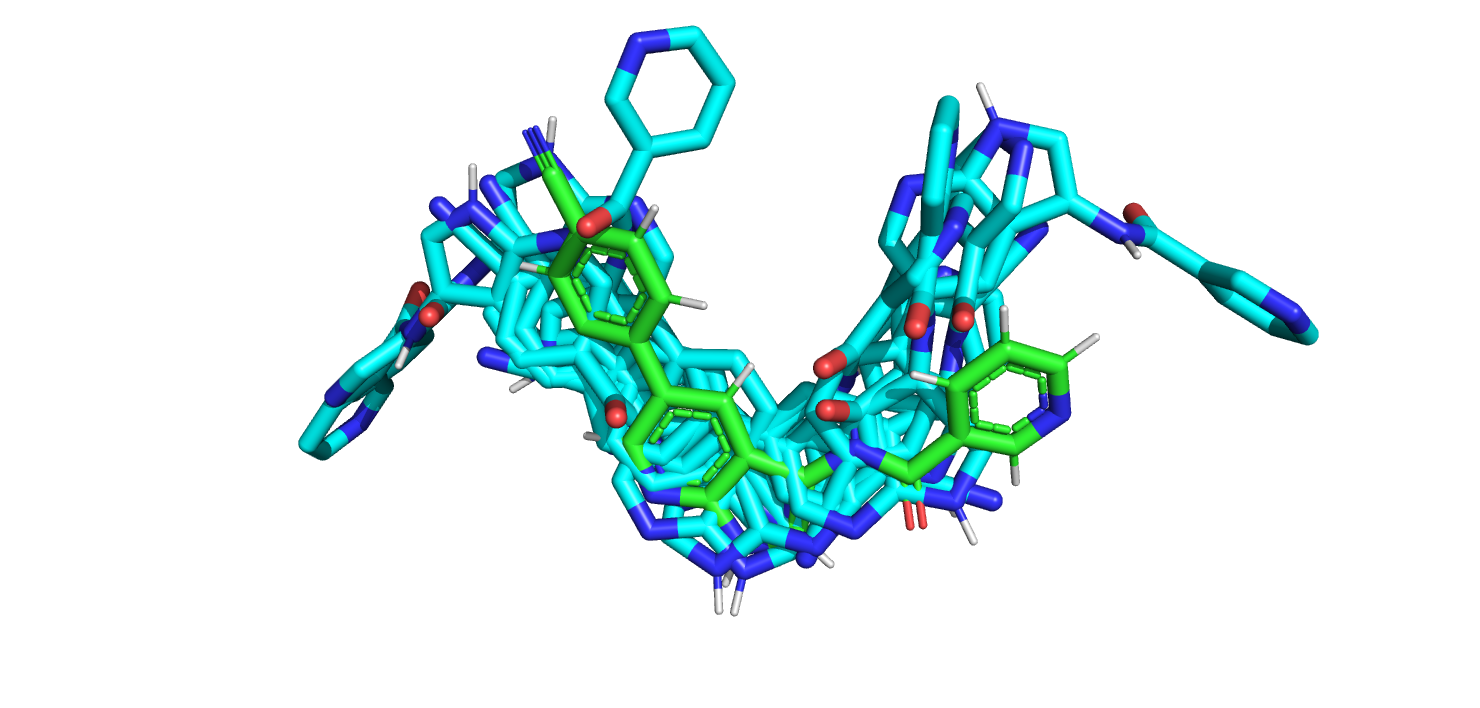
Докинг производился с помощью сервиса Webina. Были получены 9 возможных поз лиганда, изображенные на рис. 10.
Такой позы, которая бы совпадала с той, которая получена из структуры белка с лигандом. Возможно, это связано с тем, что та поза, которая
есть в связанном белке, пересекает поверхность кармана в белке, поэтому лиганд не может располагаться в двух конформациях белка в одной позе.
Рис.10. 9 поз лигандов, полученные из Webina
Рис.11. Лиганд в позе из связанной структуры и поза, первая в выдаче Webina
В выдаче Webina позы располагаются по мере убывания афинности (в ккал/моль), первая позиция соответсвует
аффинности -9.4 ккал/моль. С помощью PoseView были получены диаграммы взаимодействия лиганда и белка для позы 1 (рис.12)
и истинной позы из bound.pdb (рис. 13).
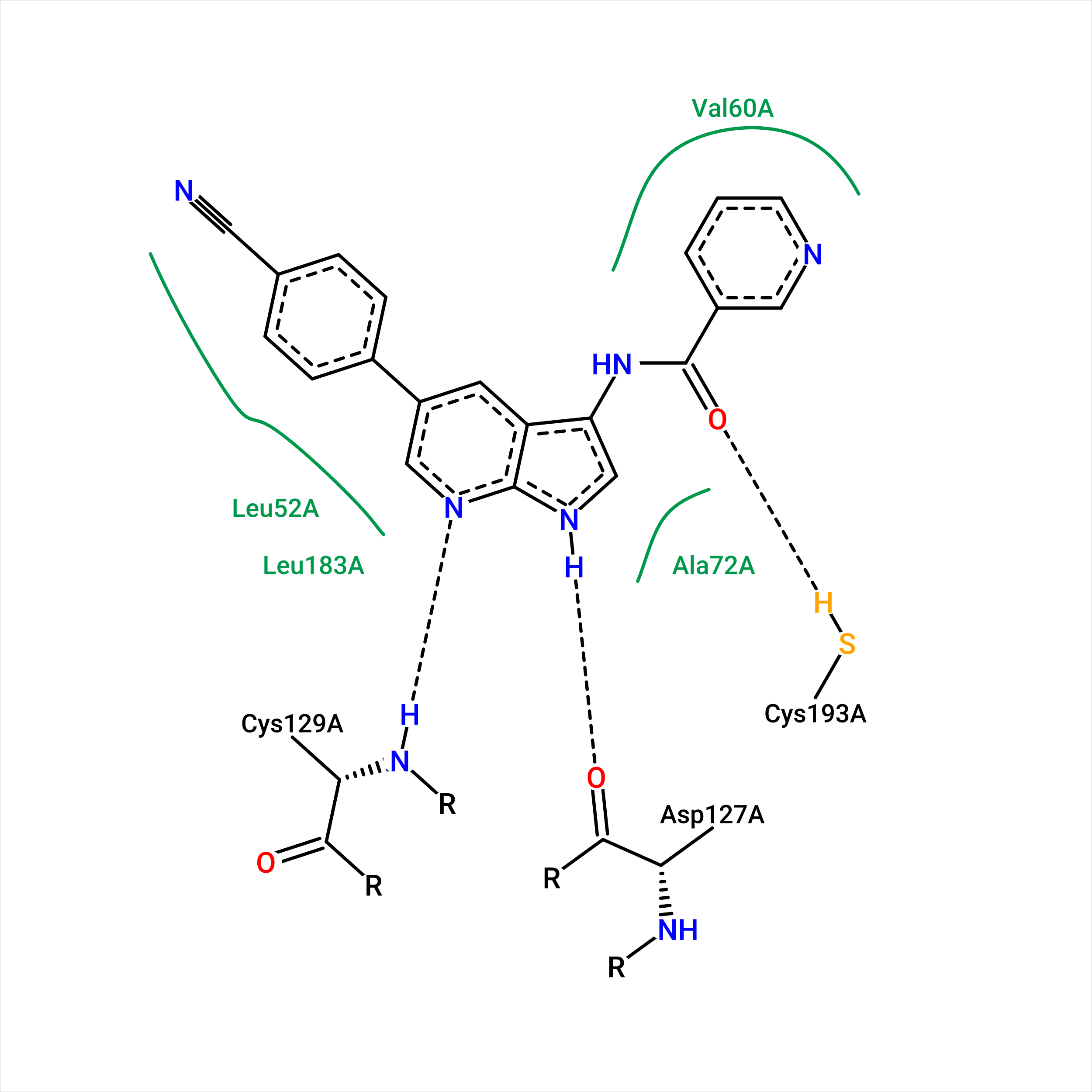
Рис.12. Диаграмма визуализации взаимодействий с лигандом в 1 позе
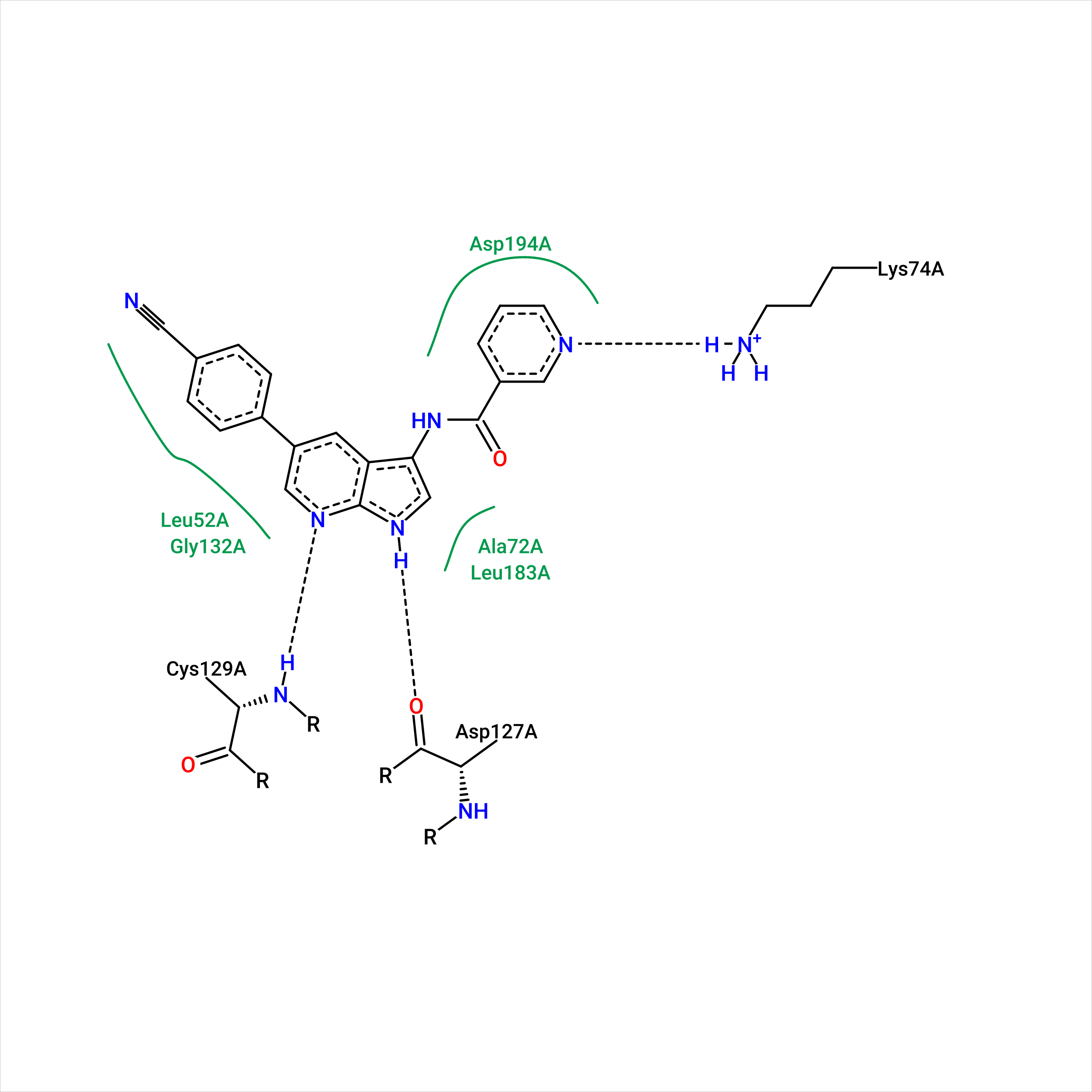
Рис.13. Диаграмма визуализации взаимодействий с лигандом в истинной позе в связанной структуре белка
Сравним полученные диаграммы. Можно заметить, что, согласно PoseView, лиганд на рис. 12 и 13 взаимодействует
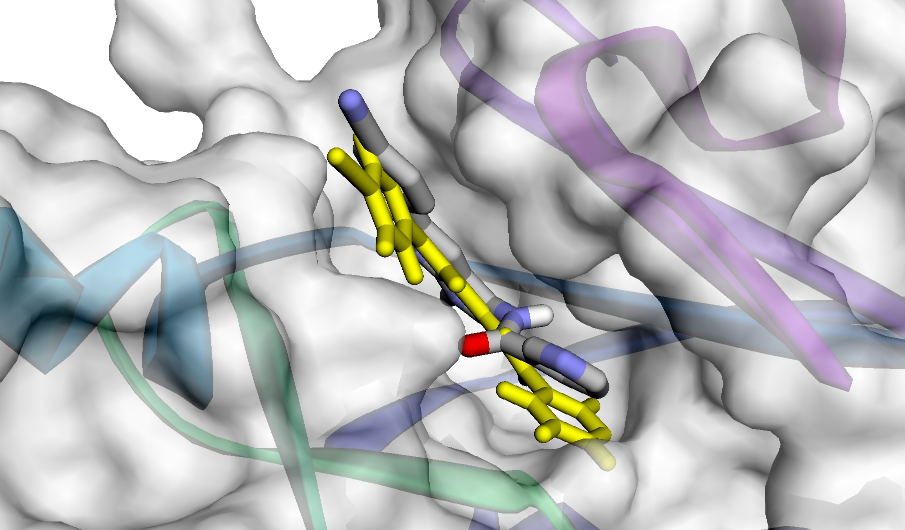
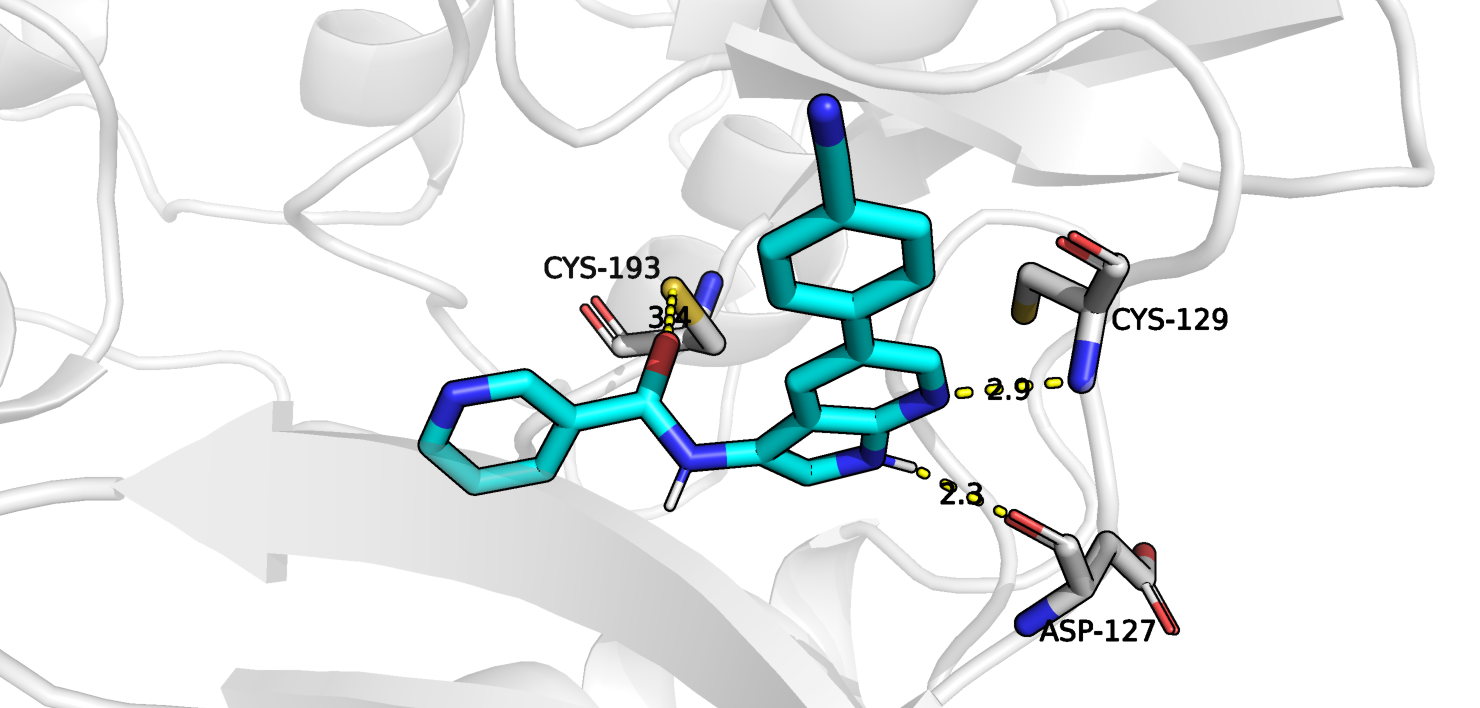
с 3 остатками белка, причем два из них совпадают и один различаются. Лиганд в позе-1, полученной при докинге, взаимодействует с остатком цистеина-129,
а в истинной позе он образует водородную связь с остатком цистеина-193.
Аспартат-197 и цистеин-129 могут быть первоначальными "якорями" в процессе связывания лиганда, задерживая
диффундирующий мимо лиганд, тем самым инициируя замыкание дополнительных связей. Примечательно, что хотя положение лигандов,
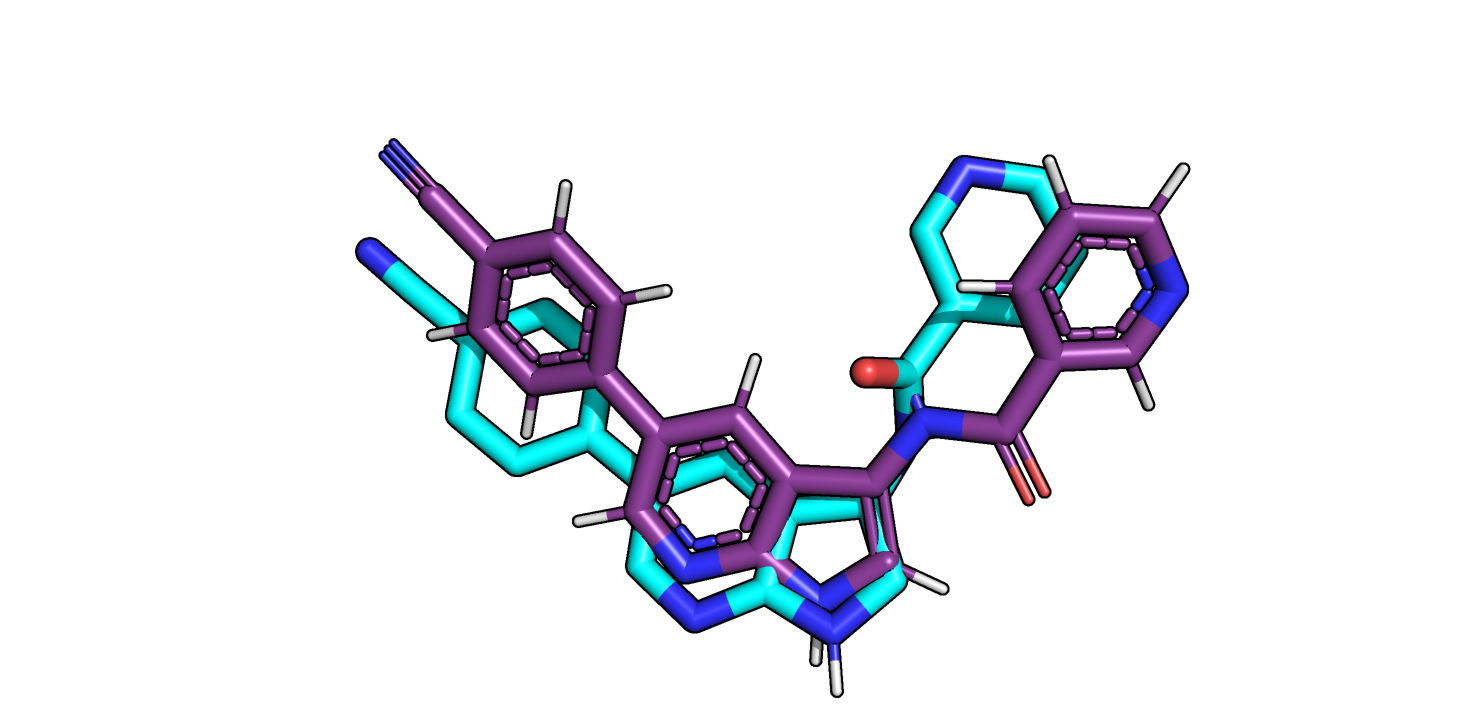
в целом, не совпадает, расположение 6 и 5 членного азотсодержащих циклов, с которыми происходит взаимодействие, почти не изменяется.
Это может означать, что индуцированного соответствия в данном случае нет,
а есть вызываемые связыванием лиганда конформационные перестройки, которые могут иметь функциональное значение.
Рис.14. Выравнивание позы 1 из free.pdb (голубая) и истинной позы из bound.pdb (розовый)
Рис.15. Поза-1 в кармане связывания, визуализация взаимодействий, показанных на диаграмме из PoseView.
Я еще немного проанализировала расположение атомов в сайтах связывания лигандов в свободной и связанной форме.
Одно из взаимодействий, которое нашел PoseView, а именно с аспартата-127, я, к сожалению, не замечала ранее. Но есть одно взаимодействие с
остатком аспарагина-136, которое, как мне кажется, может играть функционально значимую роль в процессе связывания и координации лиганда,
PoseView это взаимодействие не определил. Хотя в той позе 1, образование водородной связи между аспарагином-136 и лигандом врядли возможно, так как
расстояние между атомами 3.9 А, но в истинной позе это расстояние меньше. Интересно также, что при выполнении 1 задания я обращала внимание на
цистеин-193, но в истинной позе лиганд не может с ним взаимодействовать, а теперь оказывается, что при инициализации связвания может.