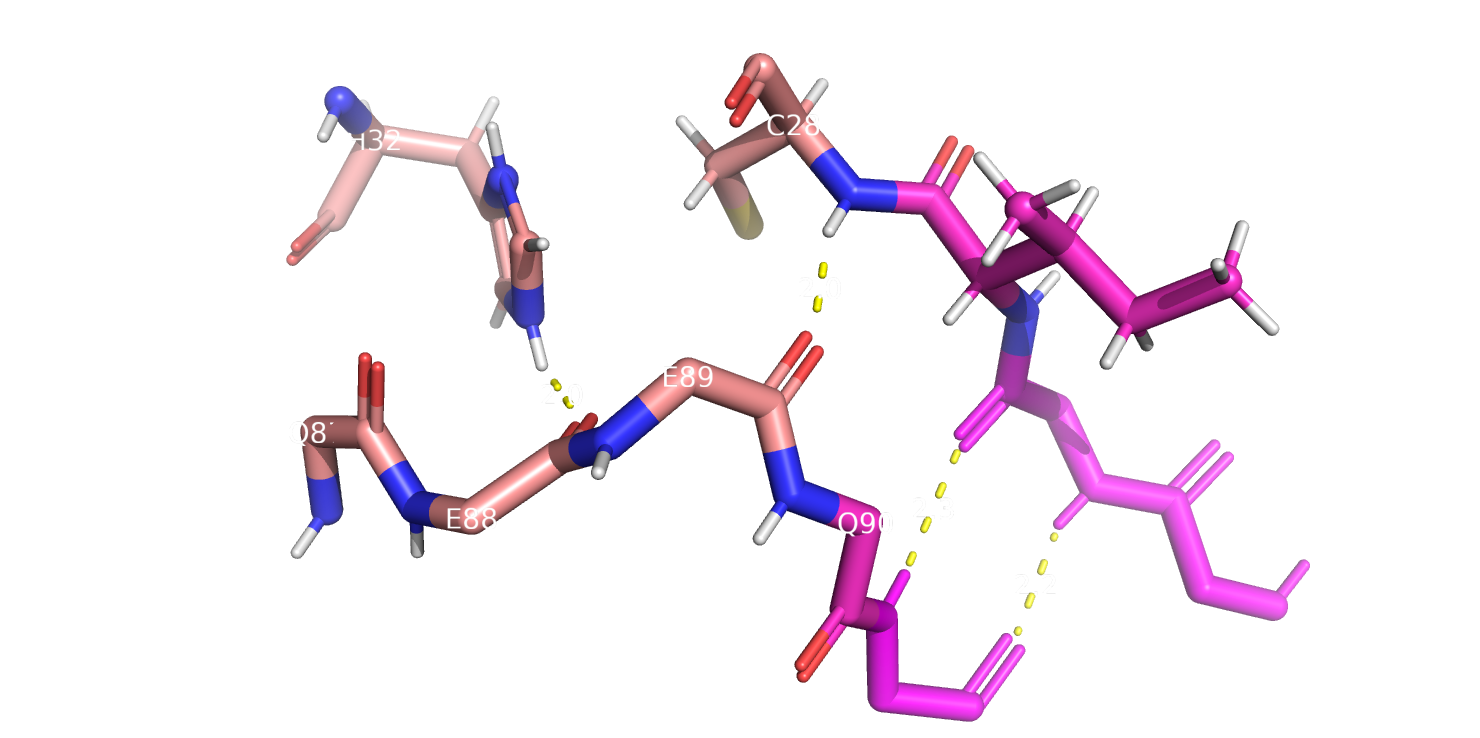Задание 1. Работа с разметкой вторичной структуры в ручном режиме
В этом практикуме я работала со структурой 2AXW из 6 практикума. С помощью 2Struc я получила аннотацию
вторичной структуры посредством использования разных алгоритмов. На рис.1 изображено выравнивание аннотации от DSSP, STRIDE, PSEA, STICKS.
Аббревиатуры в выравнивании: H = спираль, E = бета-лист, O = другие,
X = неопр./No Consensus, B = остаток в изолированном бета-мостике, S - изгиб,T = поворот с водородной связью, C - coil, неопр.
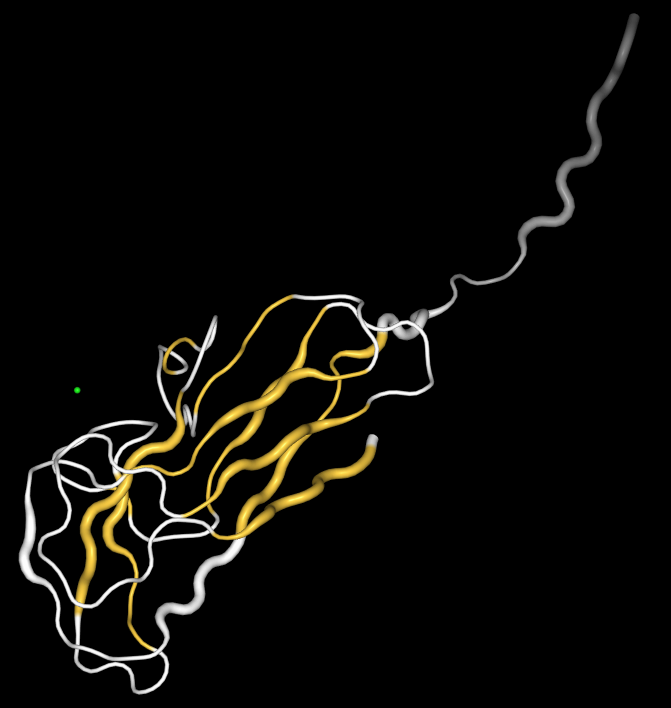
Первое, что можно сказать, данный белок - beta-белок так как не имеет альфа спиралей.
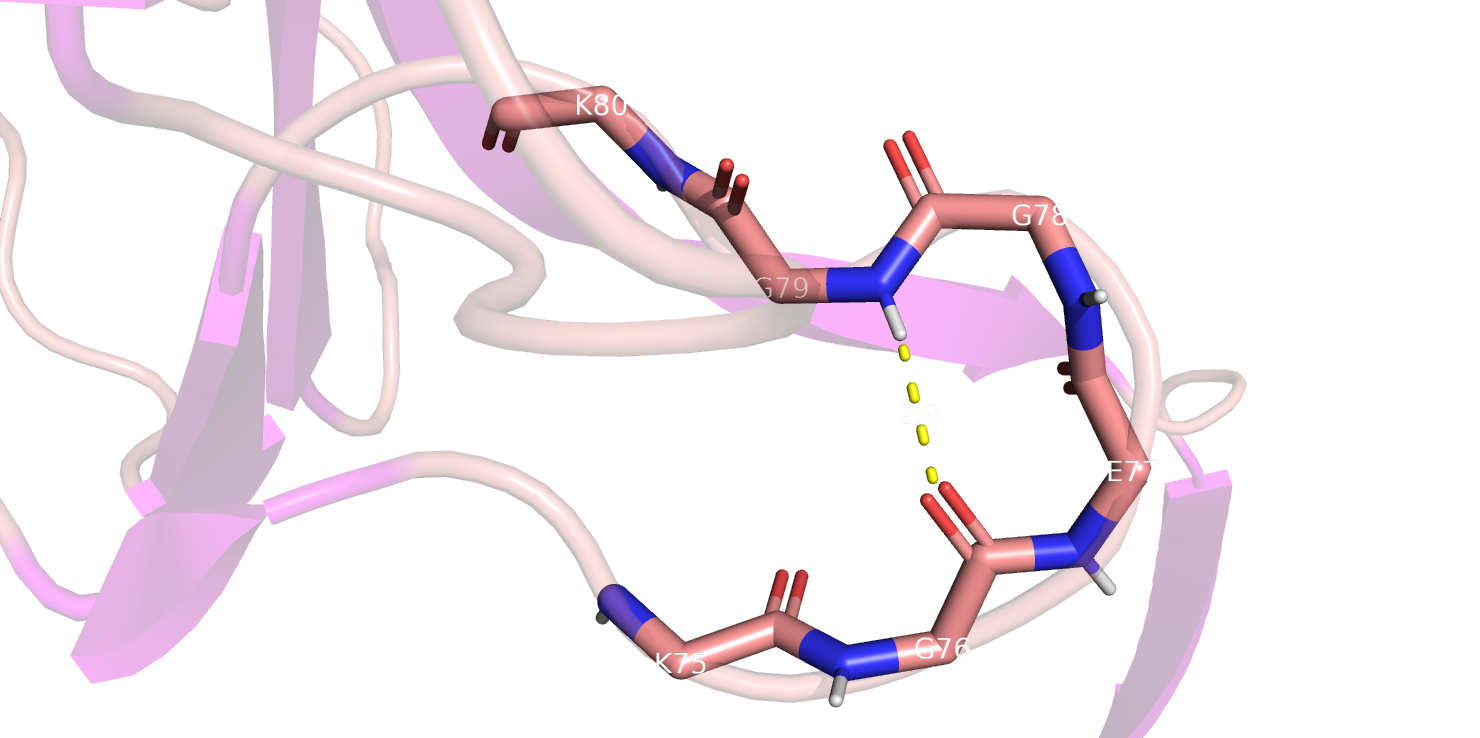
Рассмотрим позиции 76, 79. 76 позицию DSSP определил как S - изгиб, а Stride - как T. Насколько я смогла разобраться, это вообще-то не
то что разные вещи, наверное, было бы логично назвать это поворотом, т.е. turn, потому что здесь происходит именно разворот цепи в другую
сторону, который стабилизируется водородной связью 76 остатка с 79 (i, i+3). В данном случае тогда непонятно, почему согласно DSSP
76 остаток относится к изгибу, а 79 нет, он в этом случае вообще неопределен, а в Stride он тоже turn как и 76. PSEA, STICKS в случае
этих остатков просто не смогли определиться (рис. 3).
Рассмотрим теперь остатки 88-89. Как-то определить структуру в этих позициях смог только метод DSSP, и определил он их как изгиб (рис. 4).
Остаток 88 здесь взаимодействует с группой гистидина на соседней петле, и вообще, как мне кажется, тут цепь не особо изгибается, поэтому я
бы не стала очень доверять DSSP в этом случае, а 79 остаток взаимодействует с атомом азота остатка цистеина и тут можно наверно даже говорить,
что относится к бета-листу, хотя такой вид структуры тут не обнаруживается ни одним из методов. Возможно, если остов остатка изгибается,
но при этом он образует водородные связи как в бета-листе, он все равно уже не может считаться частью бета-листа, по крайней мере алгоритмы,
которые учитывают расстояние и углы между альфа атомами углерода, не распознают такое.
Рис.2 окрашивание белка согласно вторичной структуры.
Рис.3 Участок 75-80 аминокислотных остатков
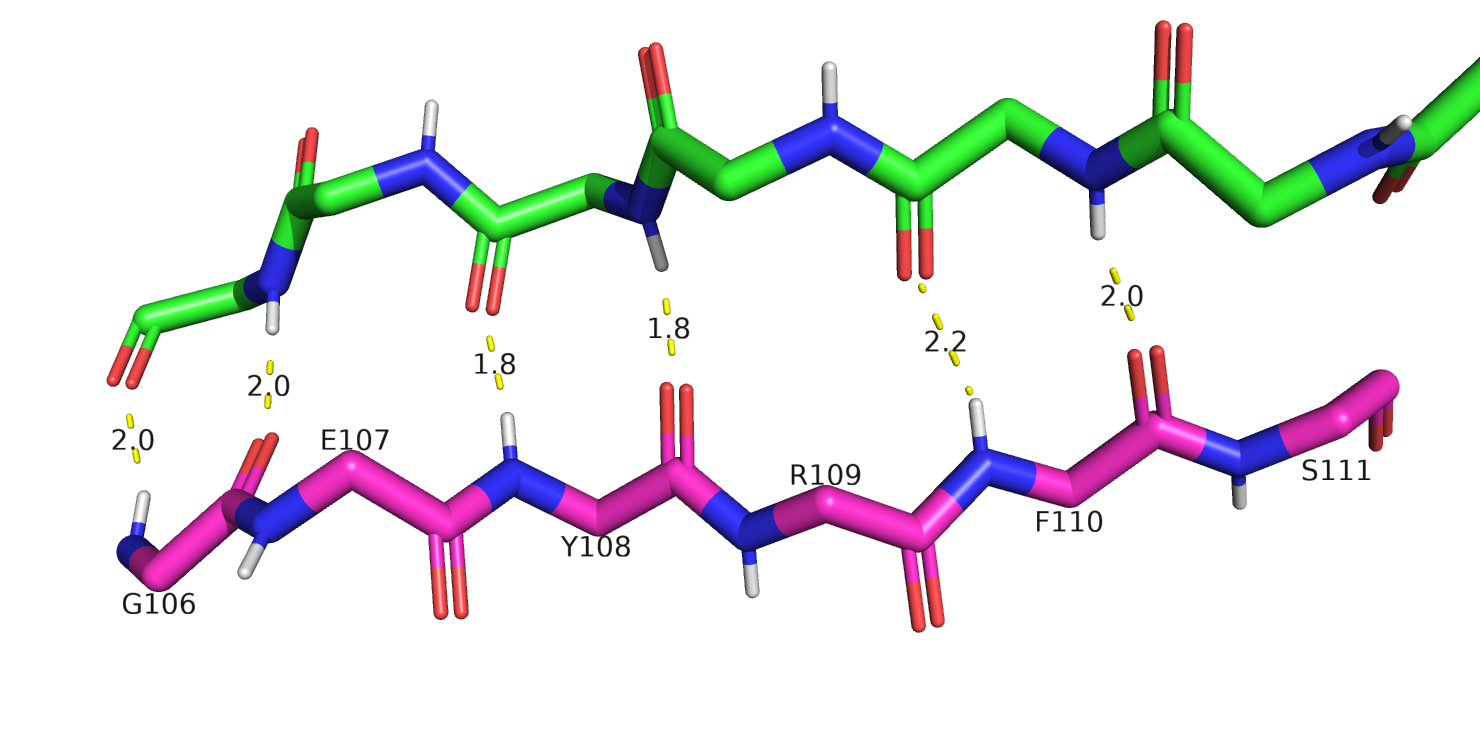
Еще один пример расхождения между приведенными методами на рис. 5 - участок бета листа в позициях 106-111:
здесь DSSP и Stride определили участок как бета-лист, а два других метода - никак не определили, хотя бета-структура тут довольно очевидная.
Возможно, все эти различия в работе алгоритмов связаны именно с торсионными углами в остове.
Рис.4 Участок, включающий 88-89 аминокислотные остатки
Рис.5 Участок, включающий 106-111 аминокислотные остатки и соседняя цепь в бета тяже (зеленая) из другой цепи белка - B
Задание 2. Работа с разметкой вторичной структуры в автоматическом режиме
Мне было выдано 30 pdb файлов. С помощью комманды " for f in /srv/www/htdocs/FBB/year_18/sbinf2021/2021.T8/
olga.shigal/*.pdb; do let "a+=1"; dssp -i ${f} -o ${a}.dssp; done;"
я получила 30 dssp файлов.
Далее, для того, чтобы посчтитать склонности каждого типа аминокислоты
образовывать тот или иной тип вторичной структуры (amino acid secondary structure propensity) я написала кое-что в python. Этот
скрипт
использует файл, в котором собрана информация из предыдущих 30 файлов, полученный
этим скриптом.
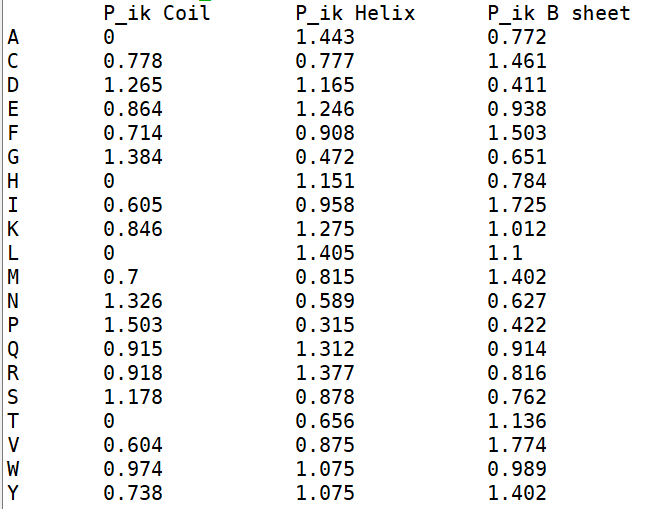
Рис.6. Таблица с аминокислотами по типу вторичной структуры
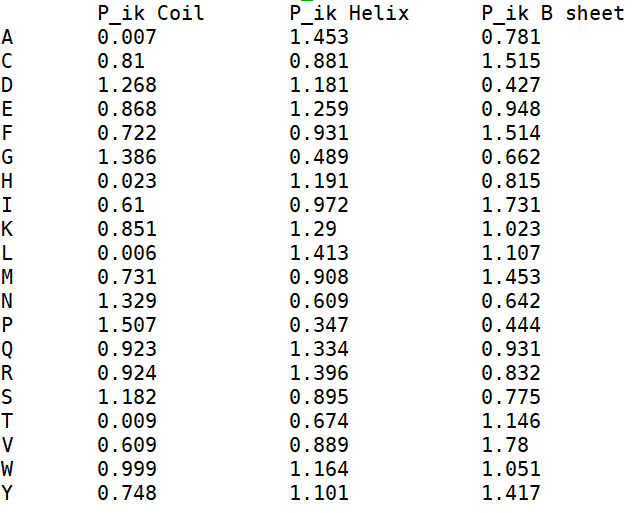
Рис.7. Таблица с аминокислотами по типу вторичной структуры с добавленными псевдокаунтами
Из таблицы на рис.6 можно заметить, что в петлях для некоторых остатков (аспартат, лизин, треонин) склонности находиться тут нулевые.
Поэтому я добавила псевдоукаунты = 1 (рис.7), чтобы избавиться от нулевых значений n_ik, т.е.
количества остатков i в датасете, образующих тип вторичной структуры j.
Возможно, можно было подобрать другое значение, но я не очень знаю как его подбирать.
Рассмотрим таблицу на рис. 7. Можно заметить, что заряженные аминоксилоты Q, R и A чаще находятся в спиралях, чем в других классах вторичных структур. Причем,
значения для глутамина и аргинина достаточно близки между собой, а аспартат как раз ни разу не встретился в петлях в данном наборе белков.
По моим вычислениям валин оказался наиболее вероятен образовывать бета-струткруры, чем остальные структуры, и по сравнению с дргуими остатками, у него там
наибольшая склонность находиться. А вот пролин и глицин наиболее склонны быть в петлях, что согласуется с известным фактом, т.к. пролин слишком не гибкий
, а глицин наоборот слишком гибкий.