В этом практикуме я работала с белком, имеющим uniprot ID A0A286XXM8. Этому ID соответсвует рецептор 1 нейротензина из
морской свинки Cavia porcellus. Последовательность представленная в uniprot получена из Tremble, т.е. она получена переводом нуклеотидной последовательности,
аннотирована она плохо. У него есть 6 трансмембранных альфа-спиралей, домен - рецептор G-белка. В общем,
как я поняла, это G-связанный белок, т.е. не фермент, а рецептор.
В uniprot про функцию G-белок связанных рецепторов нейтротензина скаазано, что они взаимодействуют с
нейротензином (тридекапептидом) для инициирования изменения клеточной активности, опосредованного G-белком.
Известных катализируемых реакций нет. Похожие белки известны из организмов, например, дегу и и восточного тушканчика, т.е. из грызунов.
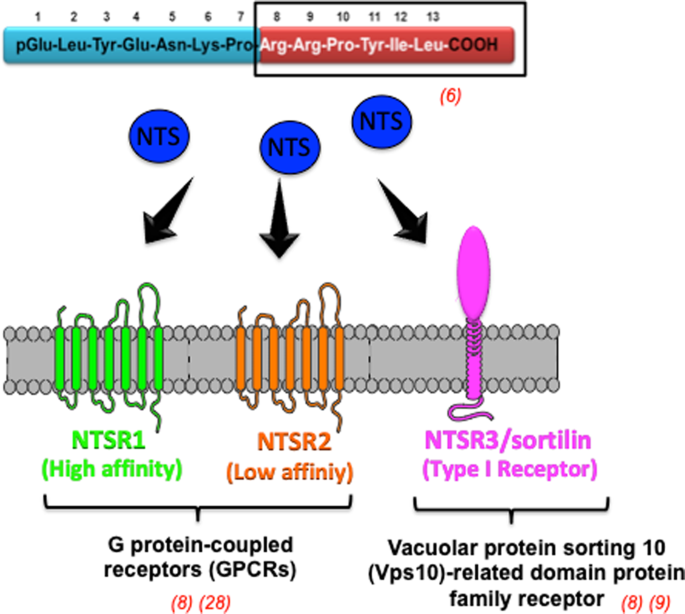
Кофакторов тоже не изветсно для данного белка. На рис. 1 представленна примерная схема взаимодействия с нейротензином
, но для человека,
и в этой схеме у рецептора трансмембранный домен из 7 альфа-спиралей, а не 6.
Рис.1.схема взаимодействия рецептора нейротензина с нейротензином .
Поиск структуры
Я сделала Protein BLAST по базе данных pdb. В выдаче получилось 100 последовательностей. Первые несколько показаны
на рис. 2. Среди находок с наибольшим скором рецептор 1 нейротензина из человека, идентичность 88.7% и покрытие 94%. Второй находкой был
комлекс аррестина-2 и рецептора 1 человека. До 14 находки включительно все структуры так или иначе относятся к рецепторам нейротензина, многие
из них получены из крыс. Немного странно, что наибольшую идентичность последовательность изучаемого белка имеет с рецепторами человека,
а не, казалось бы, родственным грызуном. Самая высокая идентичность (больше 90%) с 5й структурой в выдаче - комплекс аррестина и рецептора
нейротензина, но покрытие тут только 79%.
Рис.2 выдача BLAST
Рассмотрим первый хит из выдачи BLAST, он имеет идентификатор в pdb 6OS9_R, т.е. это R цепь
из структуры, которая называется Рецептор нейротензина 1 человека (hNTSR1) - Белковый комплекс Gi1 в канонической конформации (состояние C).
Разрешение у этой структуры 3 А, не самое лучшее. В этом комплексе несколько молекул, одна из них как раз рецептор нейротензина 1,
причем в него внесена 1 мутация в 105 позиции. Про 105 позицию было сказано в PDB, а в исходной статье я нашла, что
"Чтобы улучшить экспрессию hNTSR1, авторы сначала обрезали 19 N-концевых
аминокислот и ввели мутацию A85L". Также в комплексе есть Гуанин-нуклеотид-связывающий белок G (i) субъединица альфа-1,
Гуанин-нуклеотид-связывающий белок G (I) / G (S) / G (T) субъединица бета-1 (очень красивая структура), Гуанин-нуклеотид-связывающий белок G (I) / G (S) / G (O) субъединица гамма-2
(две небольших спирали), и молекула scFv16, получена из мыши - рекомбинантное моноклональное антитело для стабилизации GPCR/G-белок комплекса,
к естественной структуре из человека не имеет отношения, а также есть молекула JMV449 - аналог нейротензина, псевдопептид, метаболически стабильный.
Рассмотрим еще структуры из BLAST, потому что первая, конечно, хорошая, но мне кажется, ей не следует доверять полностью. Например, возьмем 2
находку в выдаче. У нее чуть-чуть выше идентичность но поменьше покрытие. ID 6PWC_R - структура комплекса аррестина-2, связанного с рецептором нейротензина 1.
В этой структуре также выделяется цепь рецептора, которая здесь без мутаций, что уже лучше. Есть молекула нейротензина, белок бета-аррестин-1,
котрый участвует в механизме передачи сигнала. Также есть легкая и тяжелая цепь белка Fab30, который узнает и стабилизирует активное состояние
аррестина. Эта структура, как и предыдущая, получена с помощью электронной микроскопии, однако у этой разрешение очень плохое - 4.9 А.
Поэтому посмотрим еще структуры.
Я посмотрела 7L0P_C, она получена из крысы, год назад, разрешение тоже 4 А, это также большой комплекс,
но именно в молекуле рецептора здесь аж 10 мутаций, это нам не подходит.
Вот кстати в структуре 4XEE нет всех этих больших белковых комплексов, есть рецептор нейротензина 1 (химерный с эндолизином), нейротензин
и несколько маленьких органических кофакторов. Тут в молекуле рецептора тоже есть мутации - 3 штуки, но меня заинтересовало здесь то,
что эти мутации располагются близко к тем трем мутациям, которые мне надо изучить в данном практикуме. А именно: мутации
E166A(3.49), L310A(6.37), F358A(7.42) изучались в статье, а мне надо изучить мутации Y145F, R317H, N344D. Это, конечно, разные по смыслу
и по расположению замены, но вдруг мне это пригодится, я пока не знаю. Разрешение у этой структуры 2.9 А, пока что лучшее.
Из этой же статьи есть еще одна структура 4XES, они очень похожи, но у этой разрешение чуть получше (2.6) и есть еще 4 мутация.
Рассмотрим еще 6UP7 комплекс нейротензинового рецептора и аррестина 2 (человеческий). В структуре есть 7 мутаций, но они относятся не к
рецептору, что радует. Разрешение здесь 4.20 А. В общем, мне кажется, можно использовать для анализа 2 и 5 находку BLAST - 6PWC и 6UP7,
так как в молекуле рецептора здесь нет мутаций.
Посмотрим на эти структуры - рис. 3 и 4.
Рис.3 6PWC комплекс, альфаспиральный рецептор слева, персикового цвета
Рис.4 6UP7 комплекс, альфаспиральный рецептор слева, розового цвета.
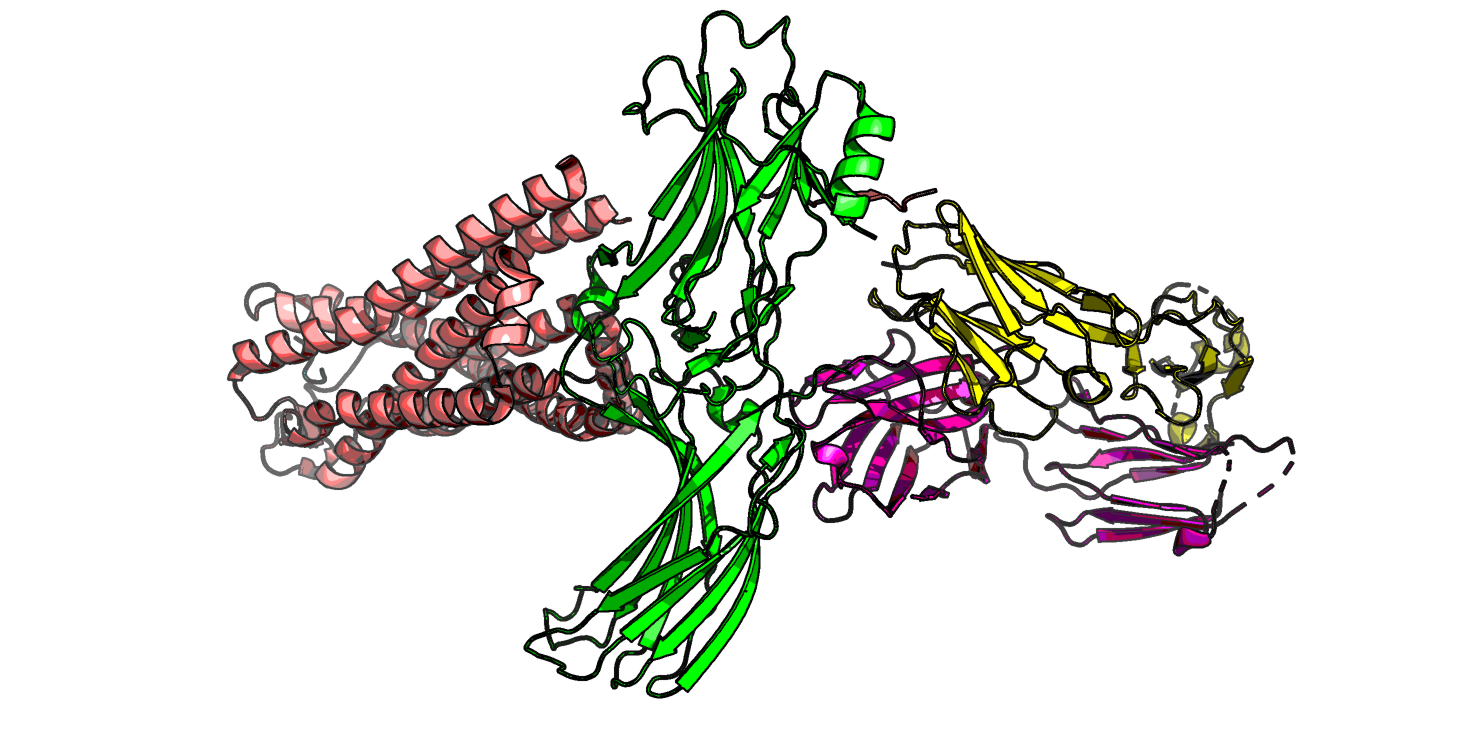
Посмотрим на структуру 6PWC для начала (рис. 5-7). На рис. 5 показано, что молекула аррестина-2 располагается
в центре альфа-спиралей рецептора нейротензина -1, со стороны, отдаленной от остального комплекса, я так понимаю, это сторона внешняя от мембраны.
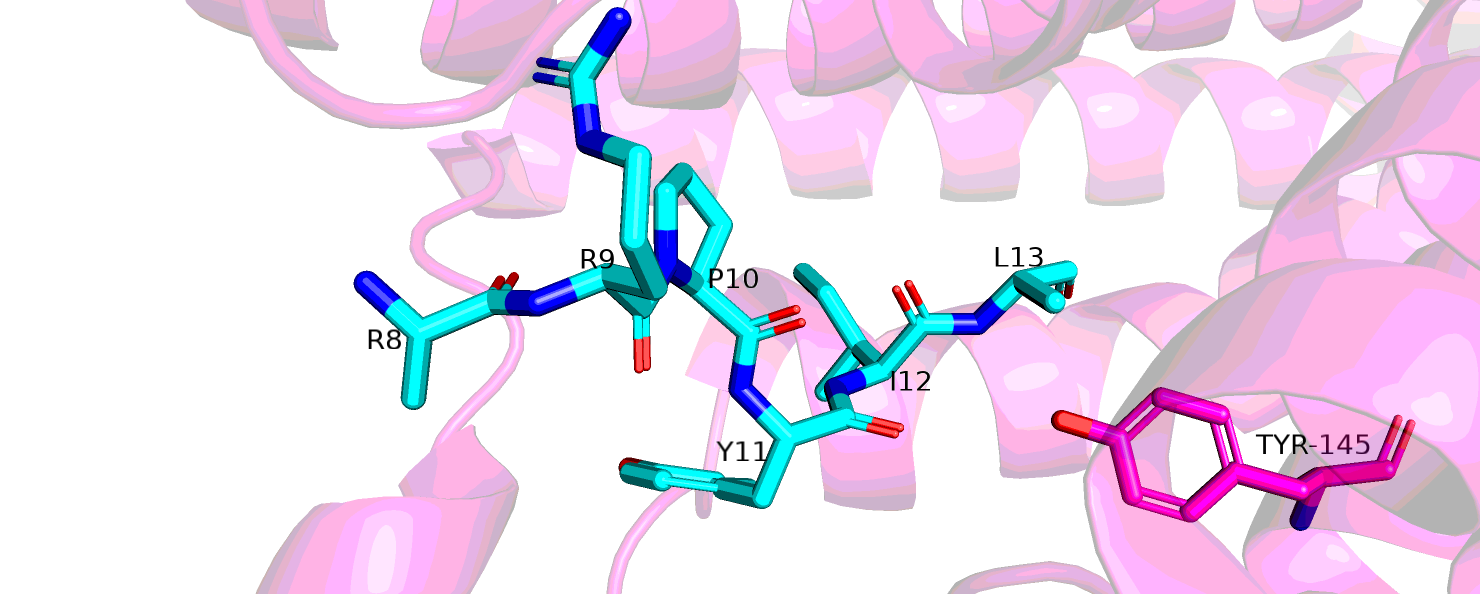
Но разрешение здесь не дает нам делать какие-то точные выводы о взаимодействии лиганда и рецептора. Нейротензнин состоит из 13 остатков, но здесь
он начинается тольько с 9, остальные не определены вообще, также радикалы очень многих остатков вокруг сайта связвания этого лиганда не определены.
Атомы остатков, которые расположены на расстоянии до 6 ангстрем до лиганда выделены на рис. 6. Видно, что из всех боковых радикалов нейротензина,
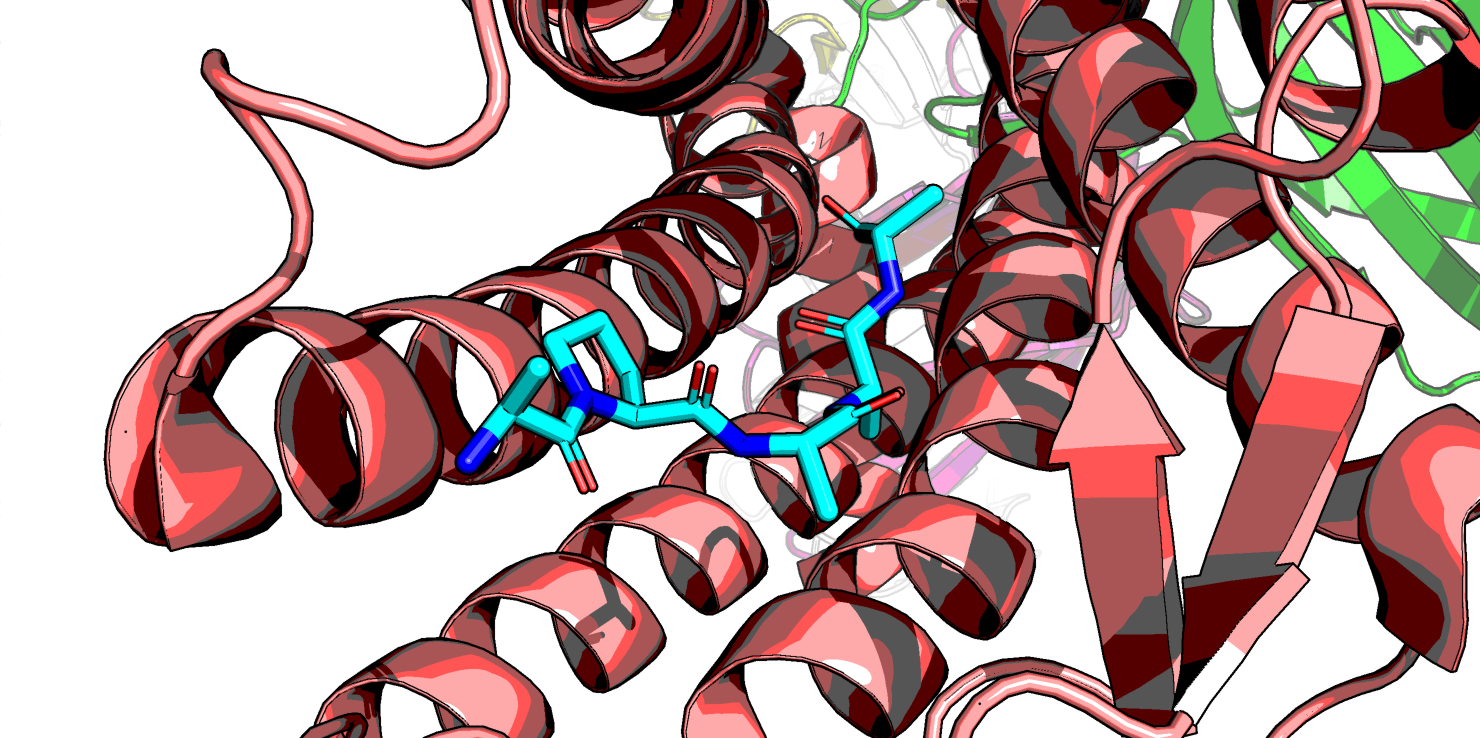
которые достаточно большие, определены в лучшем случае бета-углероды. На рис. 7 розовым выделен остаток тирозина-145, мутацию в котором
мне надо изучить. Про него можно точно сказать только то, что его полярные атомы остова взаимодействют с дргуими атомами в альфа-спирали.
А вот положение его боковой группы известно только до бета-углерода. Можно, конечно, предположить, что если продлить его в сторону
нейротензина, а боковую группу лейцина-13 нейротензина продлить в сторону тирозина, тогда, может быть, какое-то взаимодействие тут возможно, но пока не понятно.
Я не уверена, что есть смысл делать мутагенезис для данной стр-ры.
Рис.5 6PWC комплекс, рецептор в нейротензин
Рис.6 6PWC, остатки вокруг нейротензнина на расстоянии 6 А.
Рис.7 6PWC, расположение тирозина-145
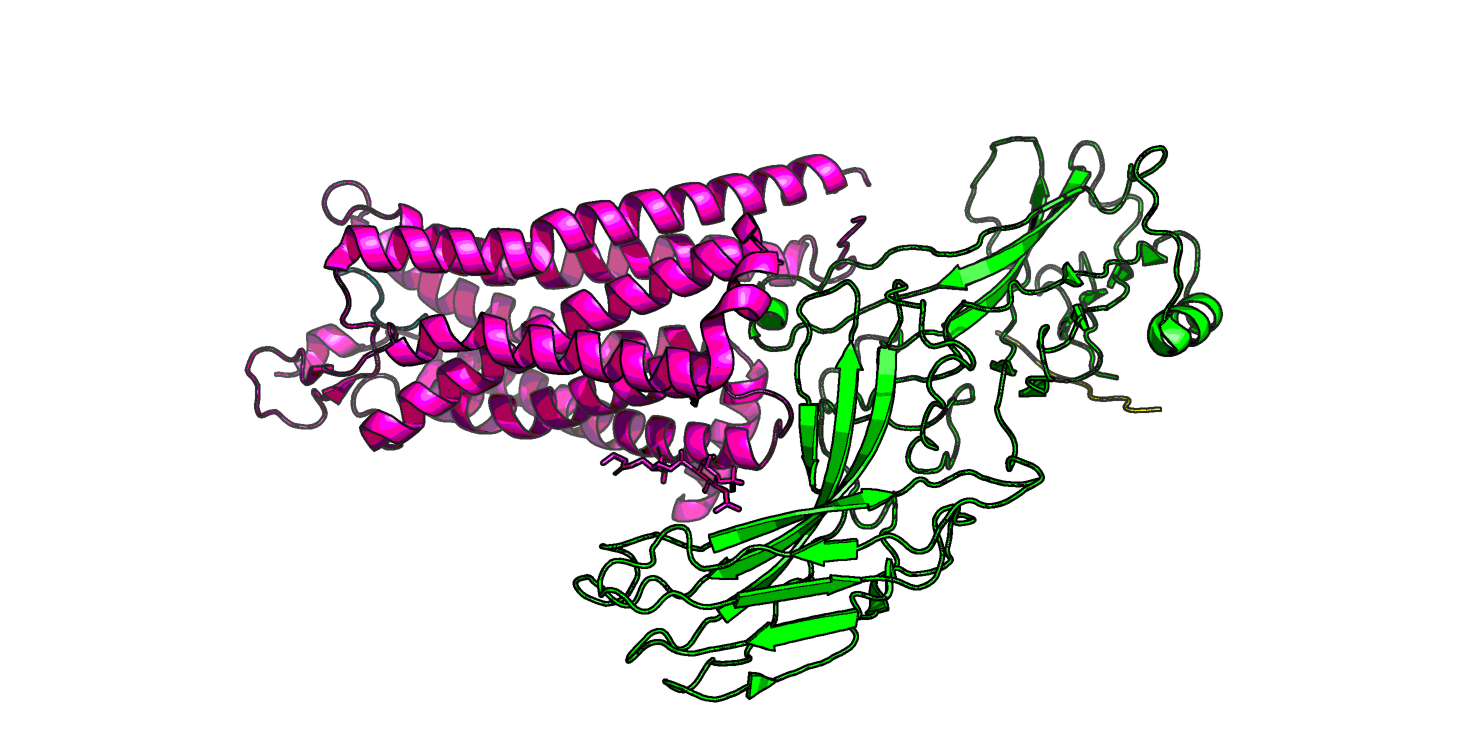
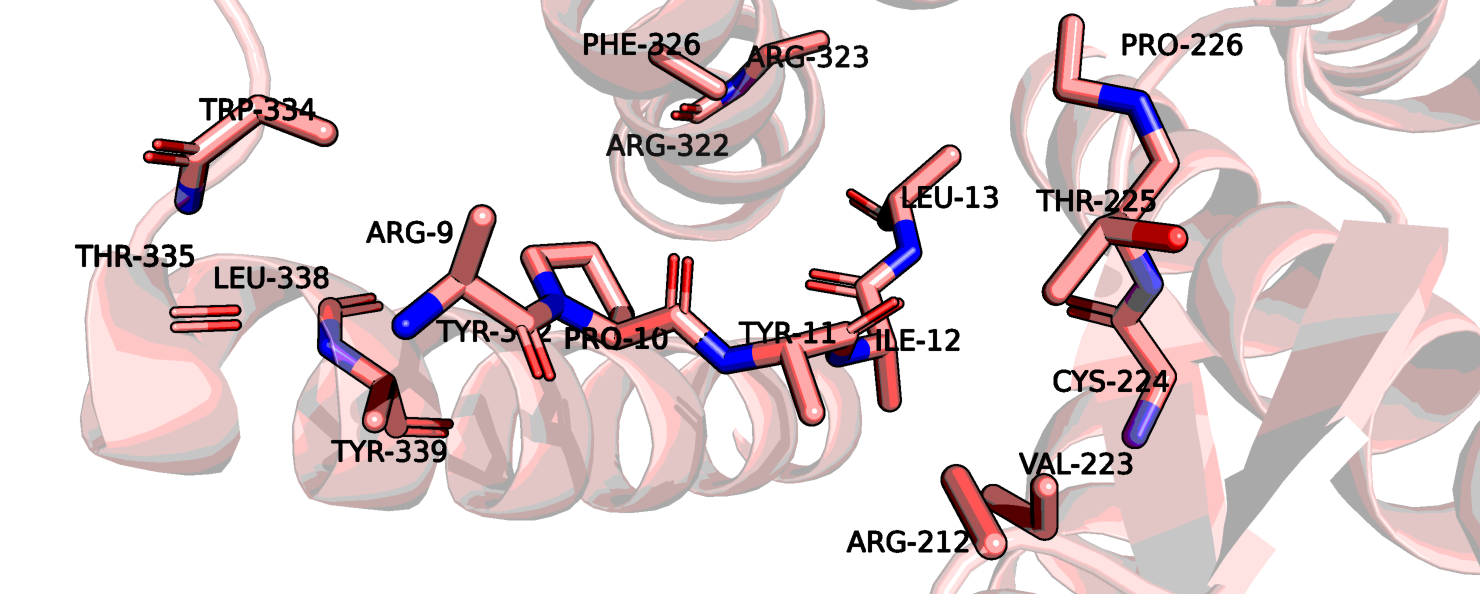
Посмотрим на вторую стр-ру: 6up7. Для нее на странице pdb написано, что в комплексе присутсвует пептид, с последовательностью,
как часть последовательности нейротензина - ARG-ARG-PRO-TYR-ILE-LEU. На рис. 8 показано выравнивание рецепторной части структур, хотя
полученные рецепторы, конечно, имеют общие участки, местами они довольно сильно отличаются. У 6PWC еще есть какой-то непонятный кусок вдали от основной цепи.
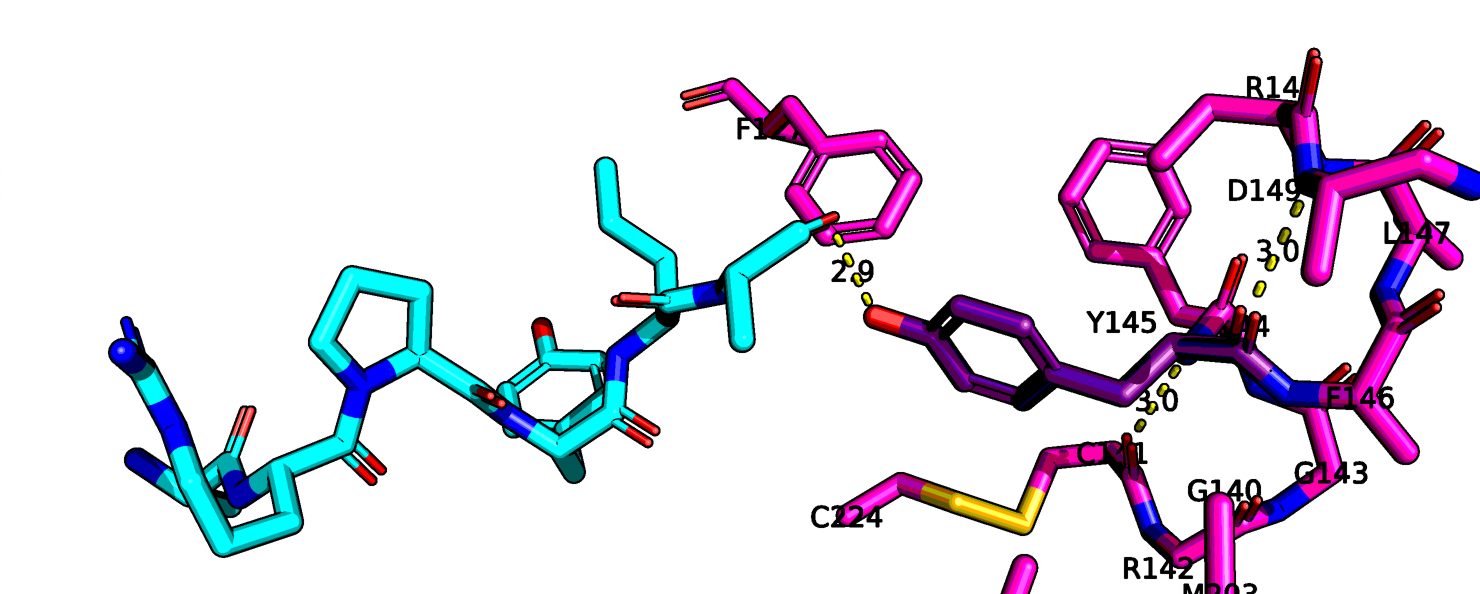
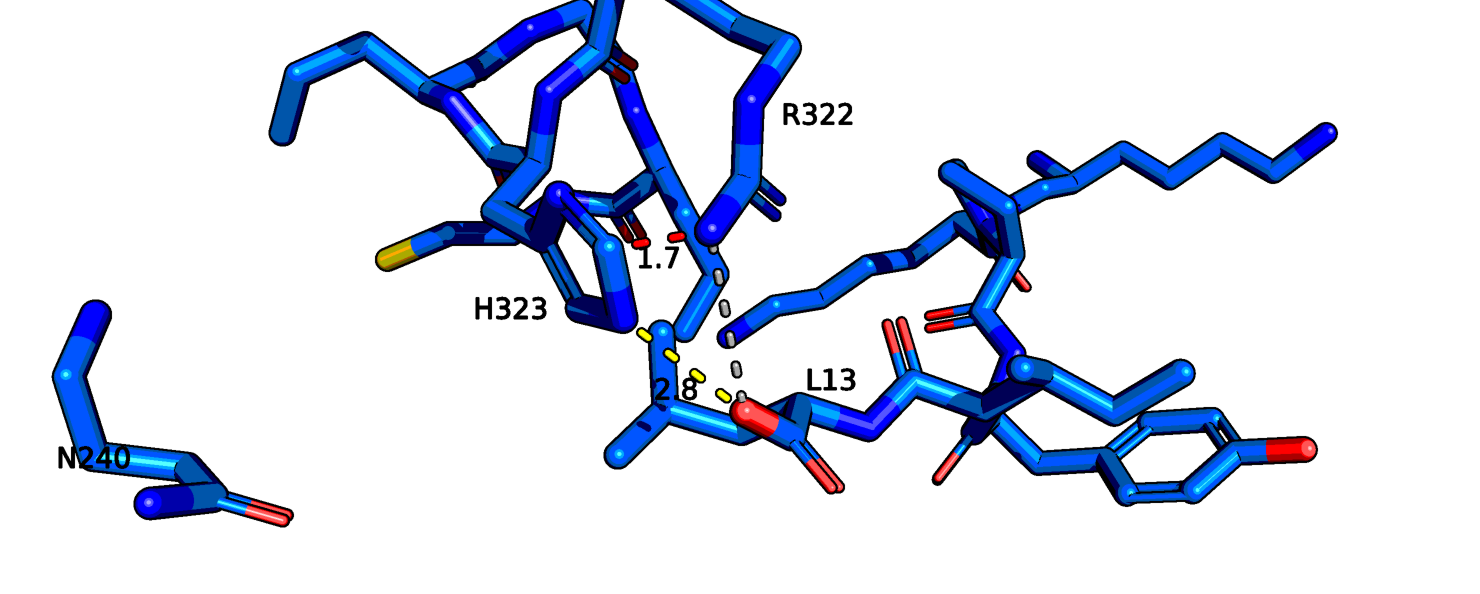
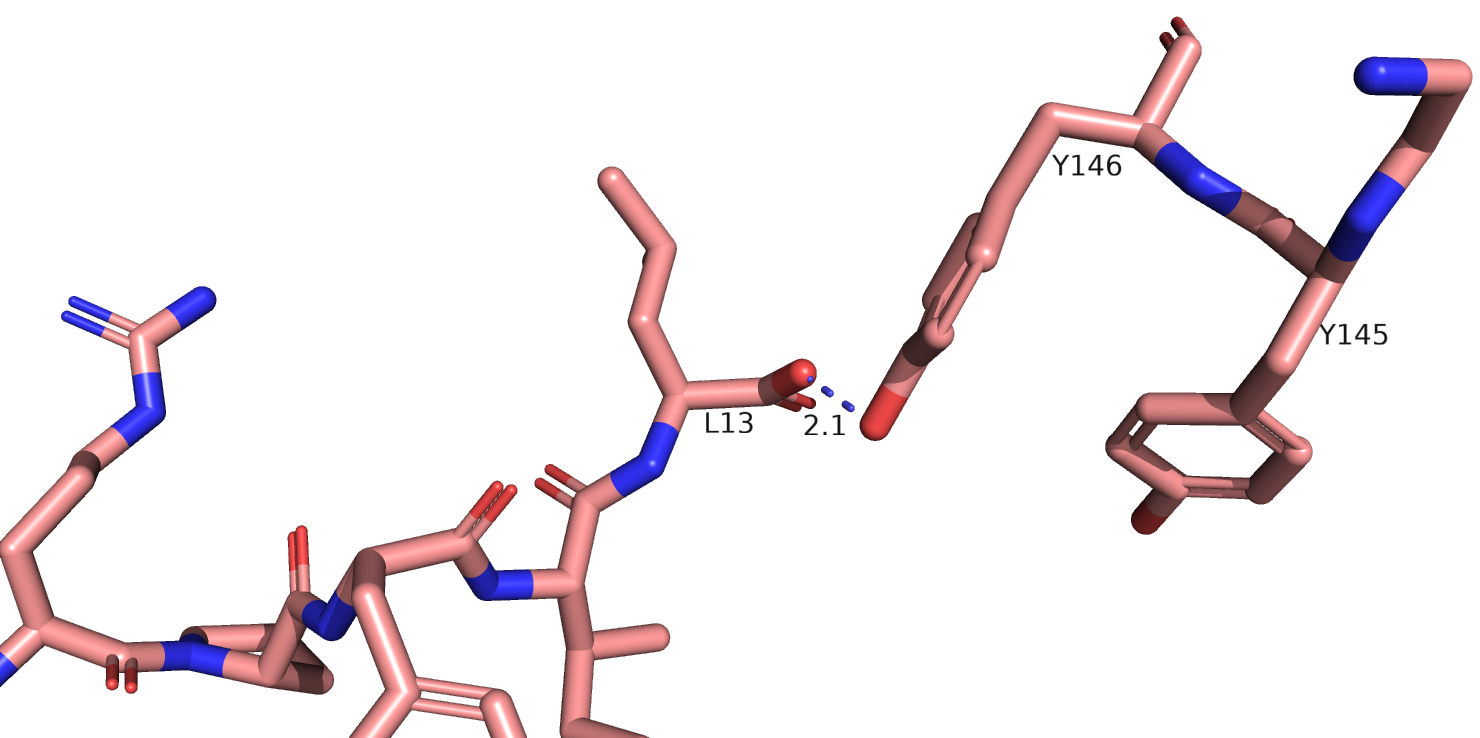
На рис. 8 я еще раз показала отдельно 6up7. На Рис.10 показан сайт связывания нейротензина. Можно заметить, что здесь лиганд определен гораздо лучше.
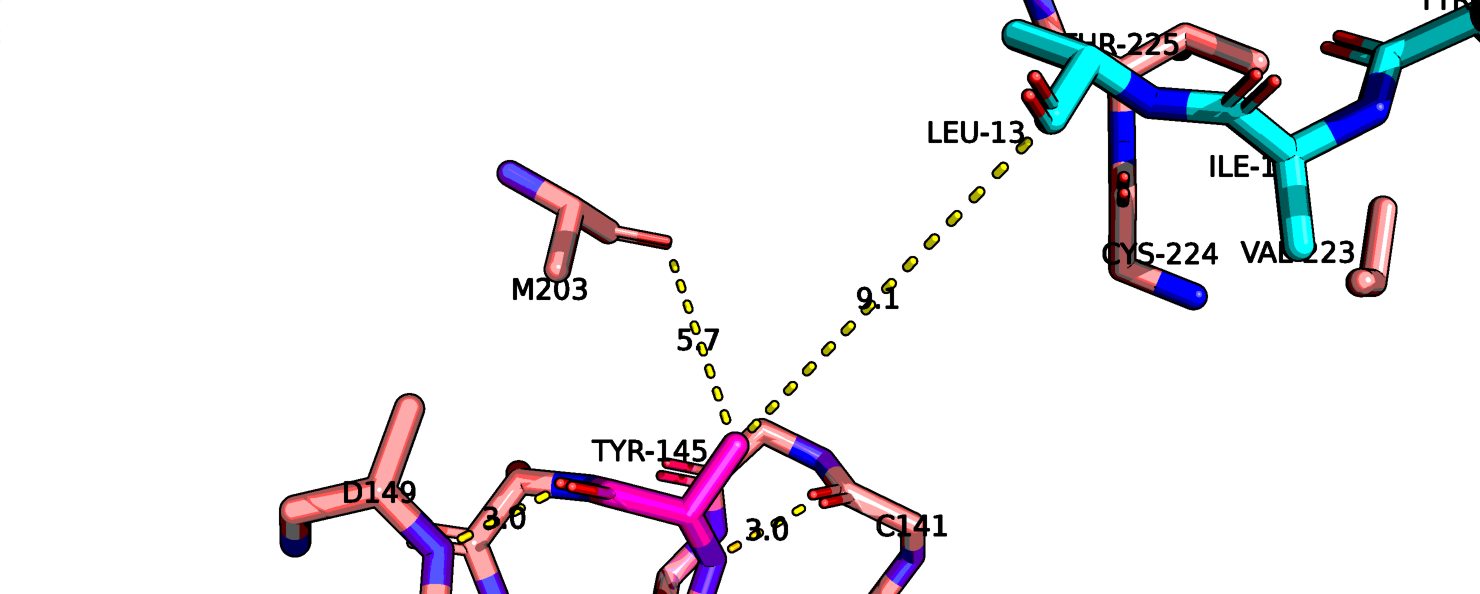
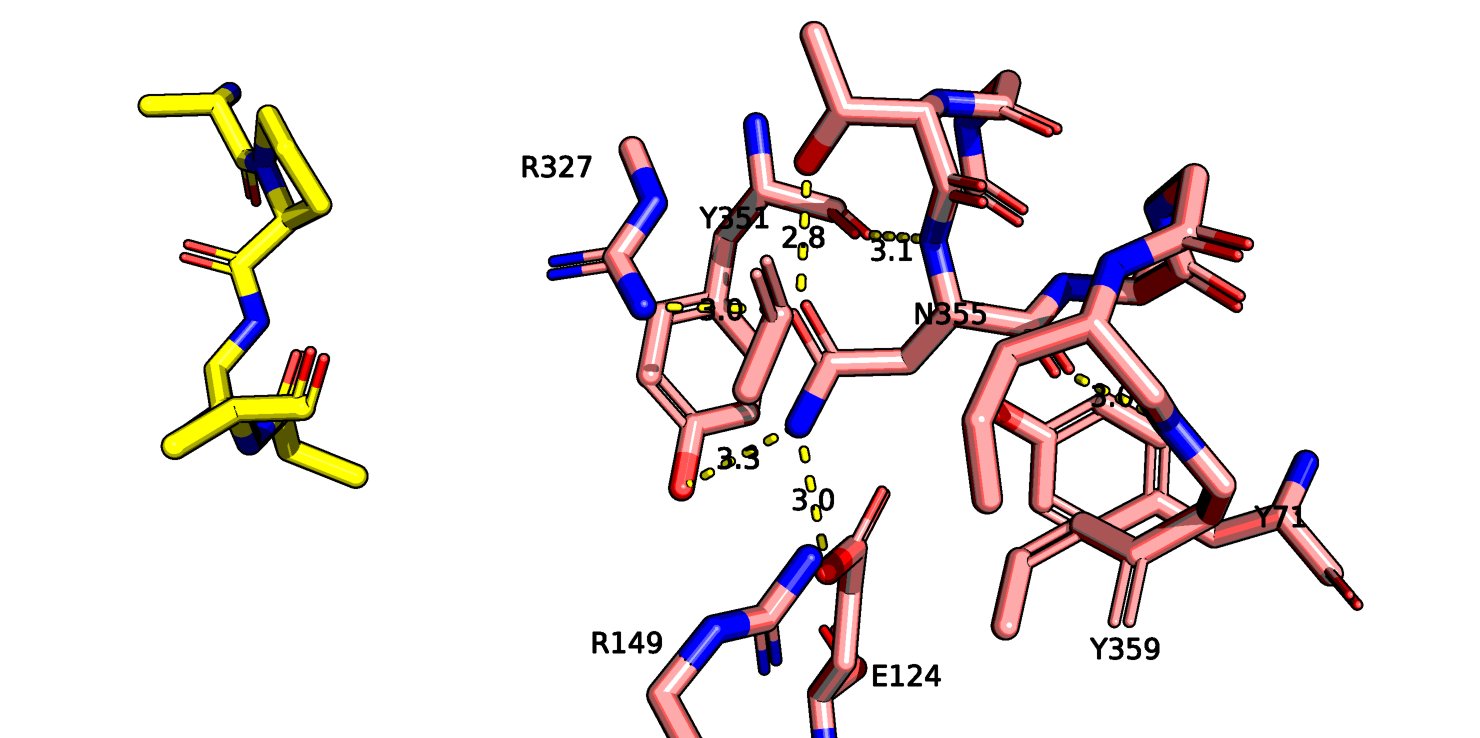
Также остаток тирозина-145 здесь хорошо виден полностью, если верить данным этой структруры, он может взаимодействовать с атомом остова лейцина-13
нейротензина. На рис. 11 показаны возможные взаимодействия этого тирозина с его окружением.
Рис.11 6up7, возможные взаимодействия тирозина-145
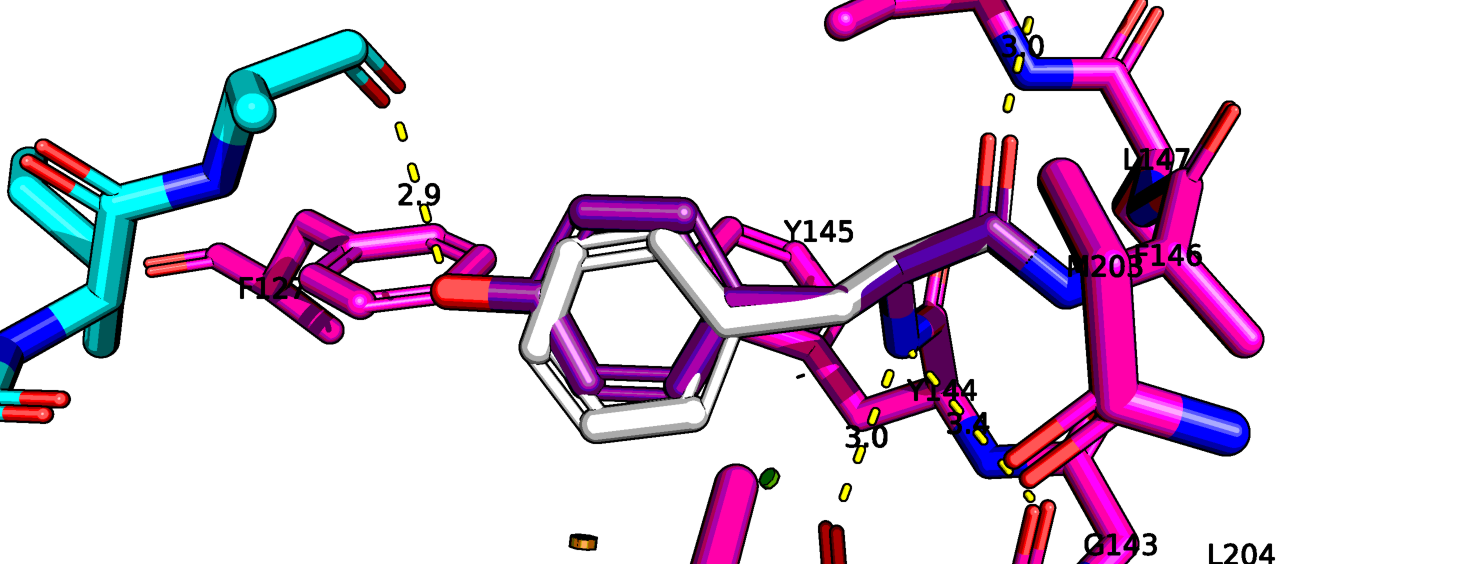
Рис.12 6up7, мутация тирозина-145 на фенилаланин (серый).
Я провела мутацию Y145F с помощью Wizard в Pymol. Изменения показаны на рис. 12. Эта функция не учитывает
изменения остова, но и без них можно преположить, что без гидроксильной группы тирозина, лейцину лиганда там больше не с кем взаимодействовать.
Поэтому можно преположить, что такая мутация (Y145F), которая ведет к ухудшению связывания лиганда, негативно скажется на функции всего комплекса.
Рассмотрим другой остаток: R-317. Он находится на одной из альфа-спиралей рецепора, рядом с сайтом связывания лиганда. В обеих используемых
мной структурах этот остаток показан только до С-бета атома, а самая интересная его часть непонятно как расположена по отношению
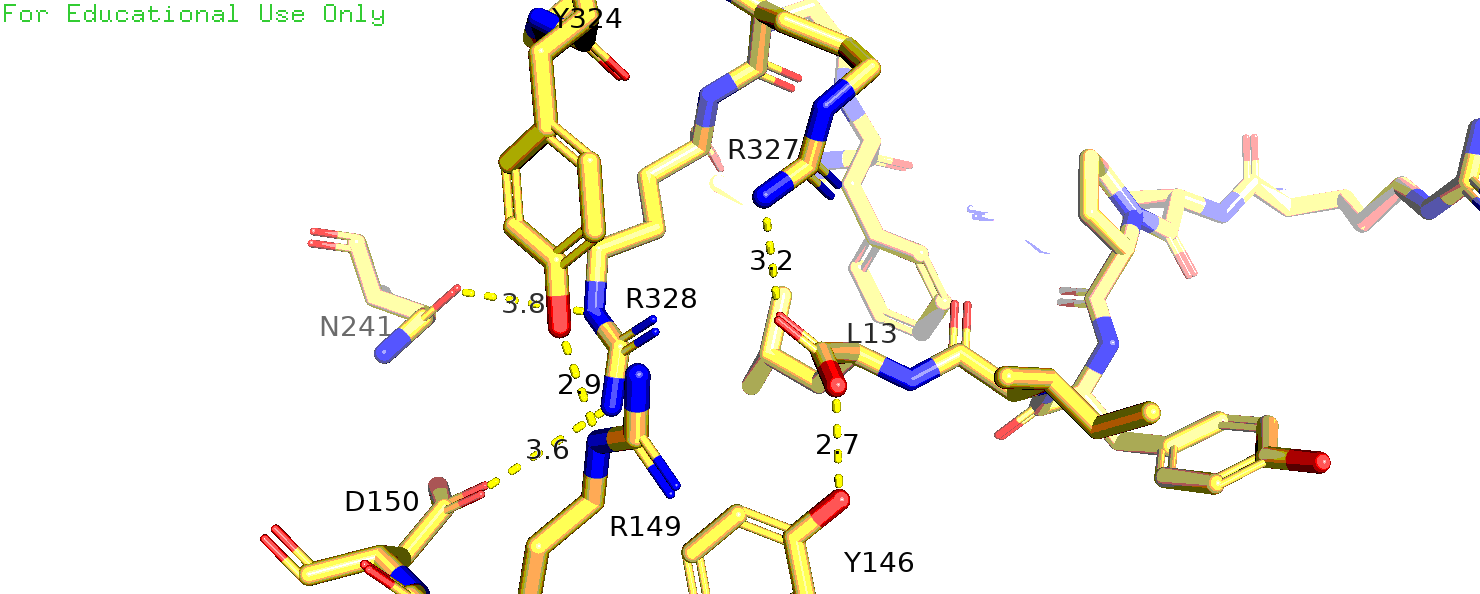
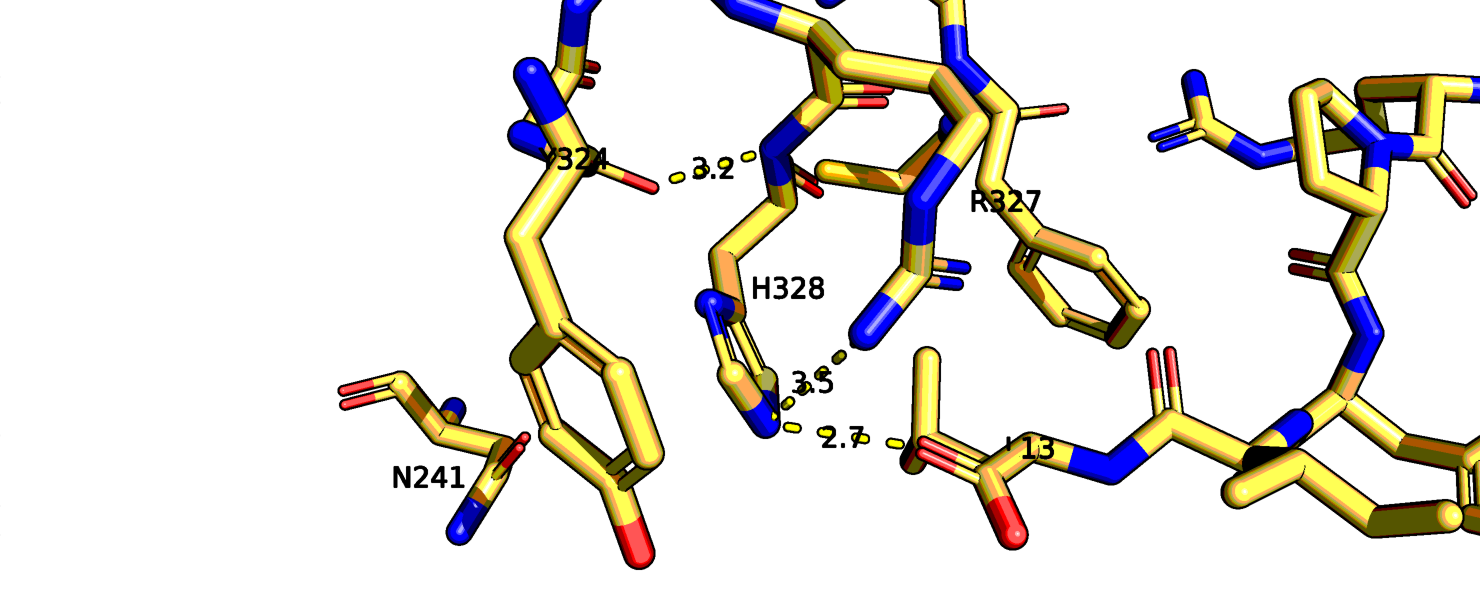
к лиганду. Я рассмотрела структуры 6os9, 4xee. В 6os9 исходный R-317 это 323 позиция, а в 4xee - 328.
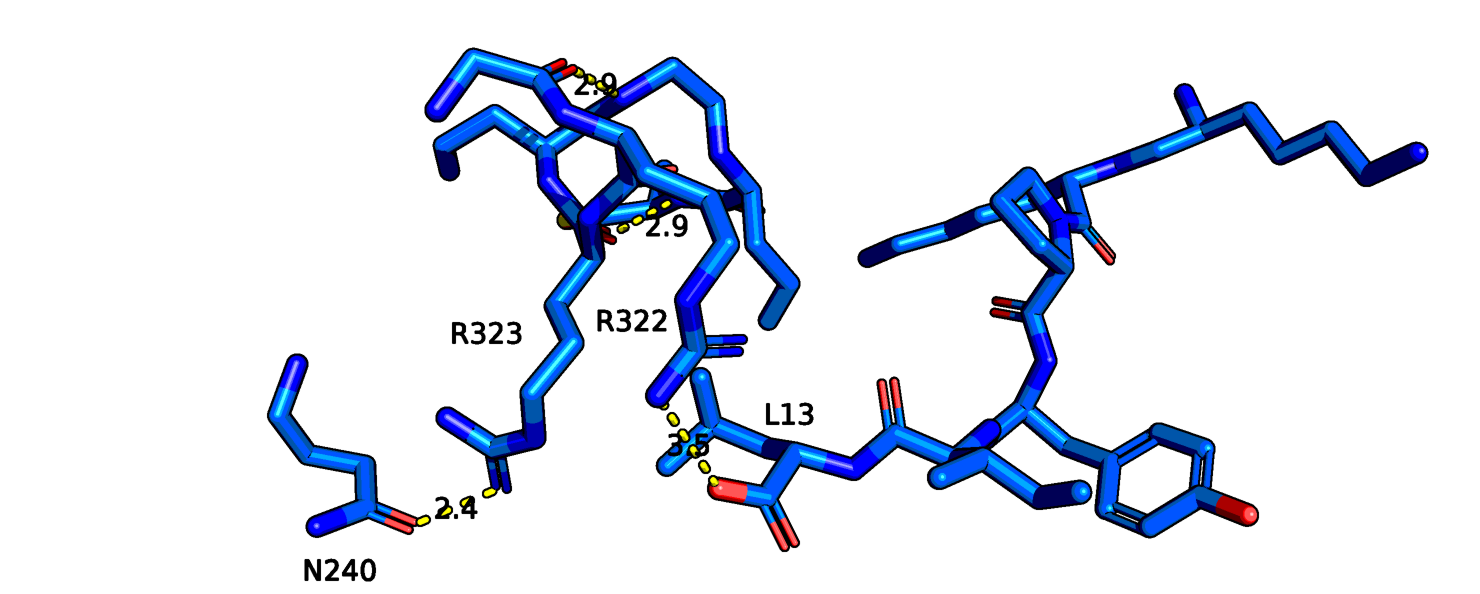
Рис.13 6os9 остаток аргинина-317(323 в PDB)
Рис.14 4xee, остаток аргинина-317(328 в PDB)
В случае 6os9 аргинин, возможно, может взаимодействовать с аспарагином-240, а соседний аргинин-322 взаимодействует с
лигандом. В целом, 6os9
больше совпадает с 6up7. Я также смутировала аргинин на гистидин (рис. 15). Появившийся в 323 позиции гистидин должен
как-то влиять на аргинин-322,
так как расстоянии между ними в исходной конформации слишком мало (красн. пунктир). Можно предположить,
что если этот аргинин, который образовывал связь с лигандом,
отодвинется дальше, то он не сможет больше связывать лиганд, но зато гистидин, который теперь расположен ближе к
лиганду, может компненсировать это.
. Возможно, это будет не очень энергетически выгодно, т.к., скорее всего, потеряется две водородные
связи - с аспарагином и аргинина. Если предположить,
что отодвинется гистидин, то связь аргинина с лигандом останется. Таким образом, судя по структуре 6os9,
мутация R317H может оказаться слегка негативной, уменьшив аффиность лиганда, т.к. возможно, связи с разными лигандами имееют разную энергию,
но все же количество связей врядли изменится.
Теперь рассмотрим структуру 4xee. В ней немного другое положение остатков, но также исследуемый аргинин взаимодействует не с лигандом, а с D150
(по нумерации 4xee), а с аспарагином, мне кажется, геометрия не позволяет взаимодействовать. Соседний аргинин тоже взаимодействует с лигандом (рис. 14).
При мутации на гистидин, расположение гистидина и аргинина такое, что они не мешают друг другу, а возможно, взаимодействуют. При
этом образуется также и новая связь между гистидином и лигандом.
Таким образом, хотя эти две структуры немного отличаются между собой, основной вывод можно сделать по ним примерно одинаковый:
замена аргинина на гистидин, возможно, является нейтральной.
Рис.15 6os9 остаток аргинина-317(323 в PDB), мутированный на гистидин.
Рис.16 4xee, остаток аргинина-317(328 в PDB), мутированный на гистидин
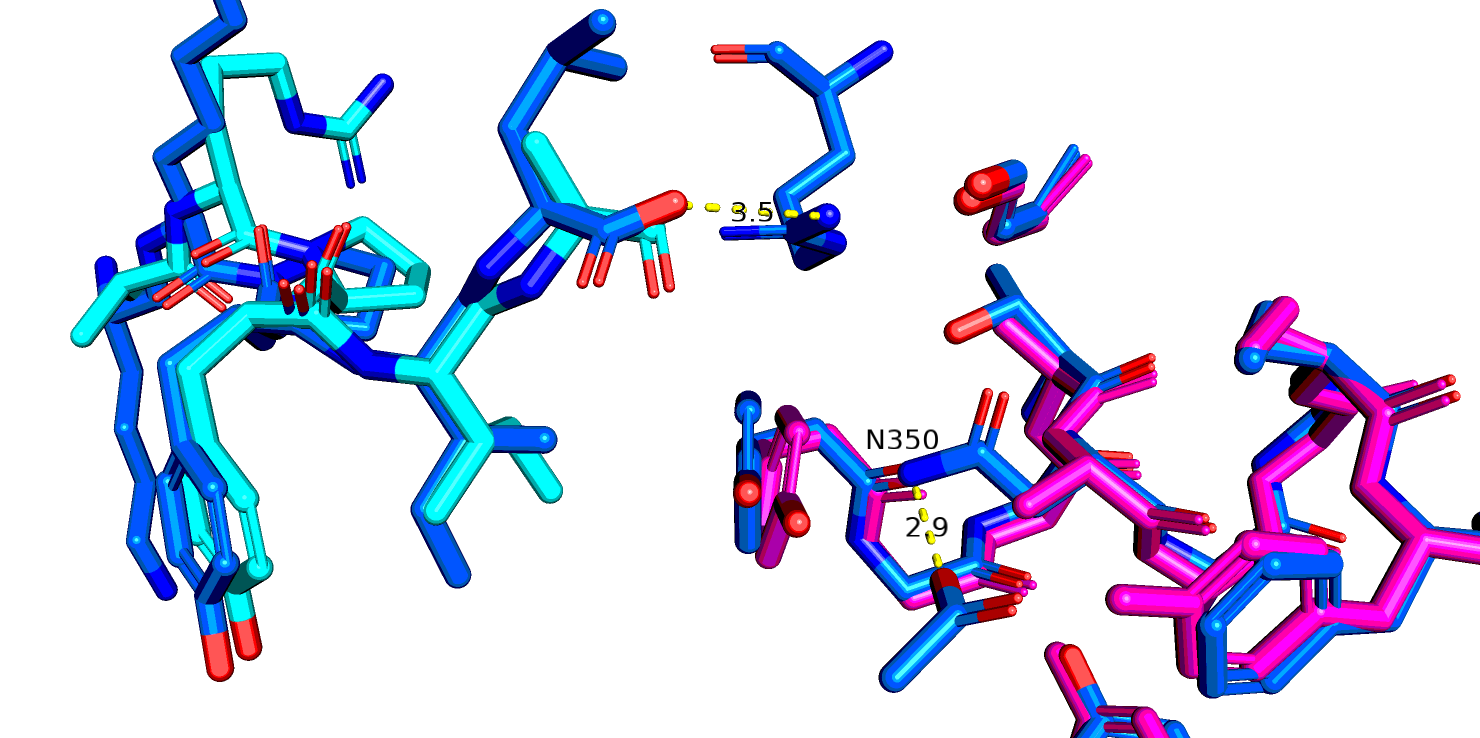
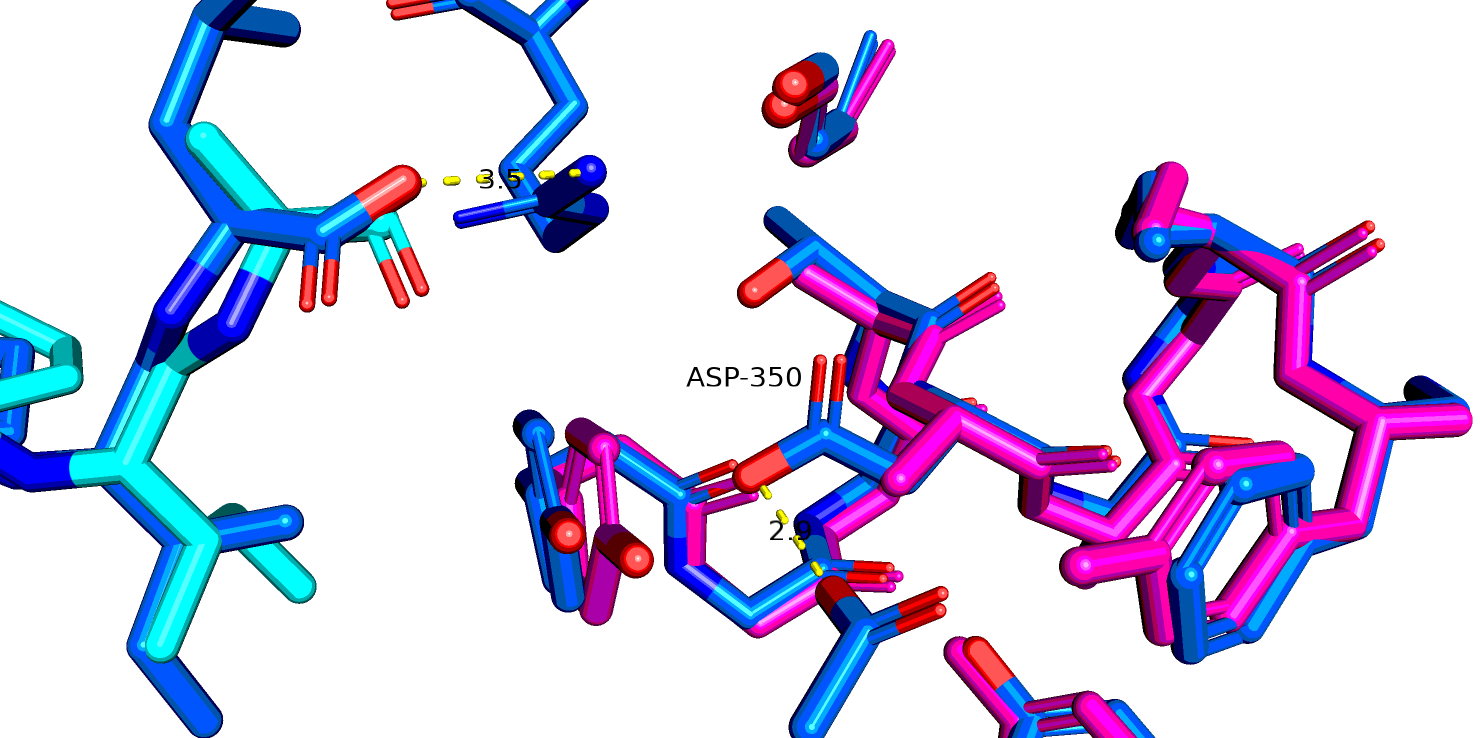
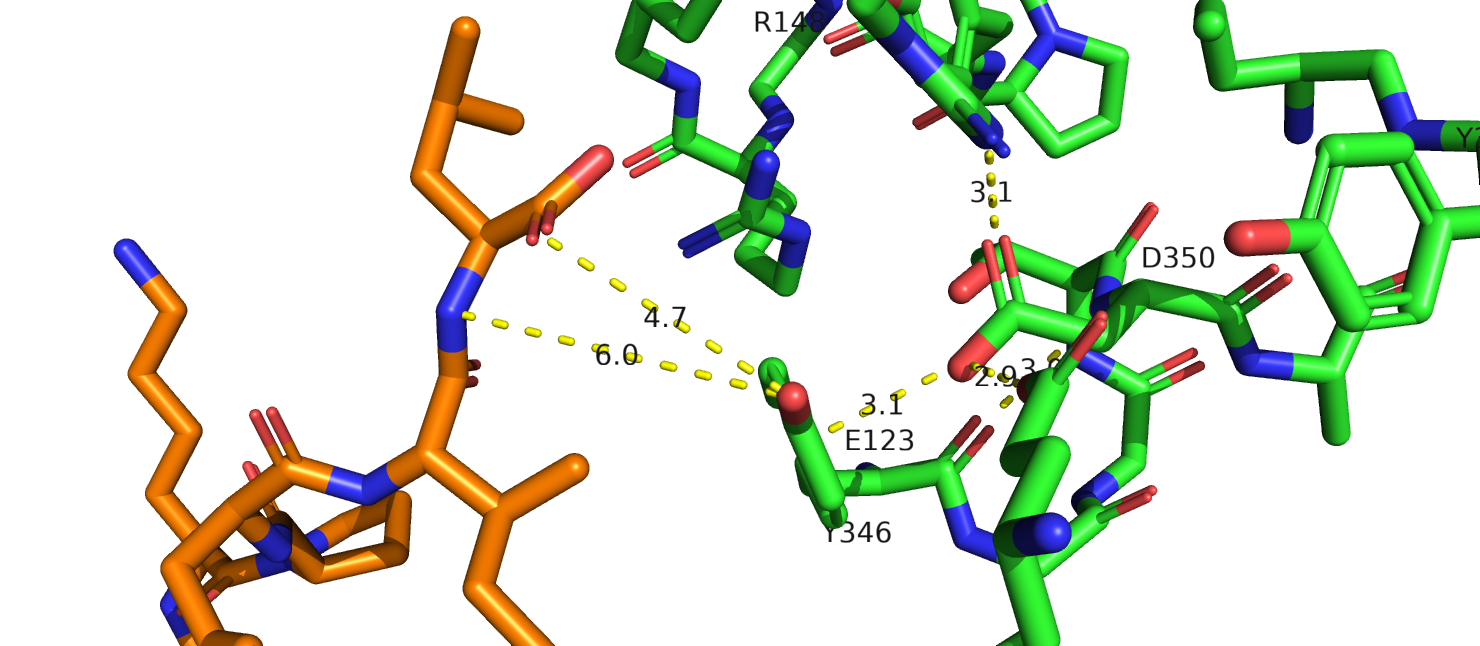
Рассмотрим еще один остаток - N344. В PDB это 350 позиция. Интересно еще, что пока я искала соответствие между Uniprot и PDB, я заметила что,
перед этим N идет последовательность MLTN в исходном белке, однако в человеческом во всех трех случаях (в принципе логично, белок то один и тот же) была
одна замена MVTN, но скорее всего эта замена никак особо не влияет на функционирование белка. На рис. 15 показано выравнивание 6up7( роз.), 6pwc (желт.), 6os9 (син.)
и выделен остаток аспарагина и лиганд из последней стр-ры. Только в 6os9 разрешение достаточно, чтобы предположить отсутствие взаимодействия между этим остатком и лигандом.
Также рассмотрела и 4xee.
Рис.18 6up7 и 6os9 (синий), остаток аспарагина-344(350 в PDB)
Рис.19 6up7 (роз.) и 6os9 (синий), остаток аспарагин-344(350 в PDB), мутированный
Рис.19 а. 4xee остаток аспарагина-344(350 в PDB)
Рис.19 б. 6os9, остаток аспарагина-344(350 в PDB)
Рис.19 б. 6os9, остаток аспарагина-344(350 в PDB), мутированный на аспартат
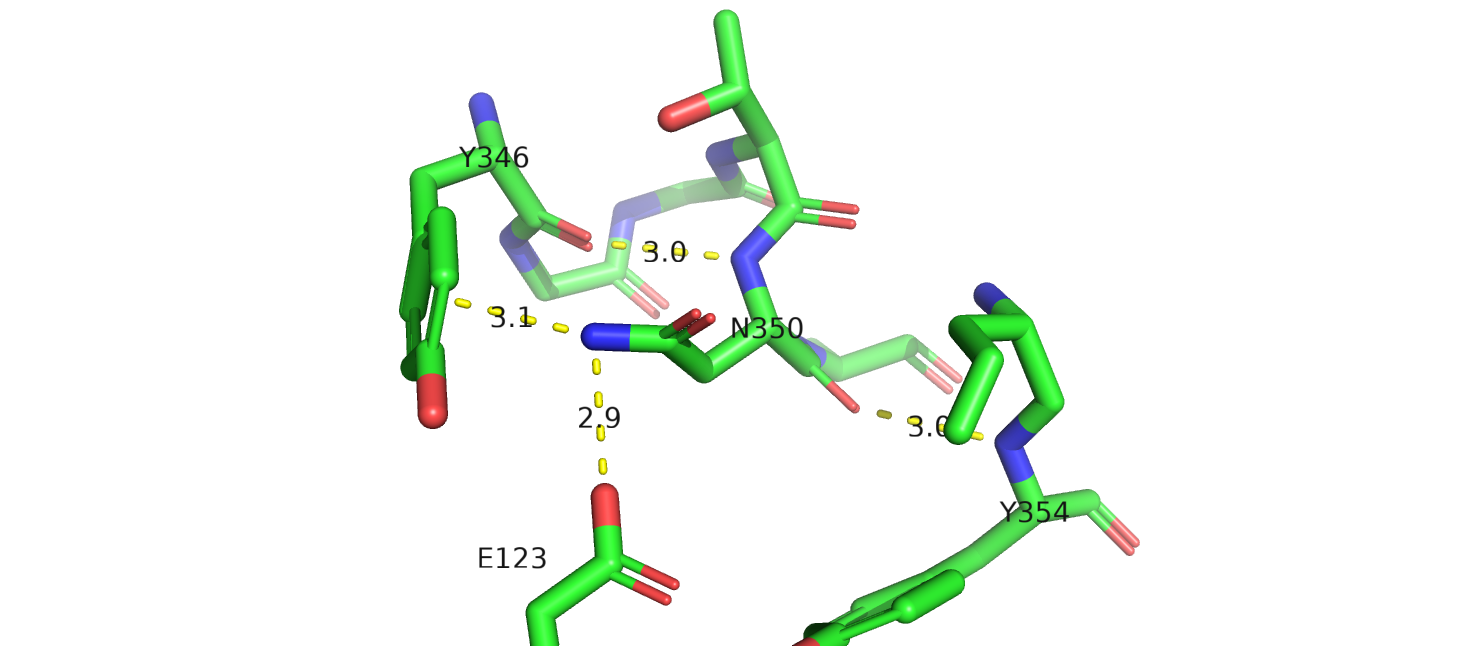
Посмотрим, с чем вообще может взаимодействовать этот остаток.В структуре 4xee (рис.19а) аспарагин взаимодействует с
глутаматом , тирозиноми аргинином. Если смутировать аспарагин на аспартат, возможно потеряется взаимодействие с гидроксилом тирозина,
эта мутация не мешает
расположению относительно соседних остатков. Кроме того, здесь нет взаимодействий с лигандом (показан желтым). Однако мутация привнеет отрицательный заряд.
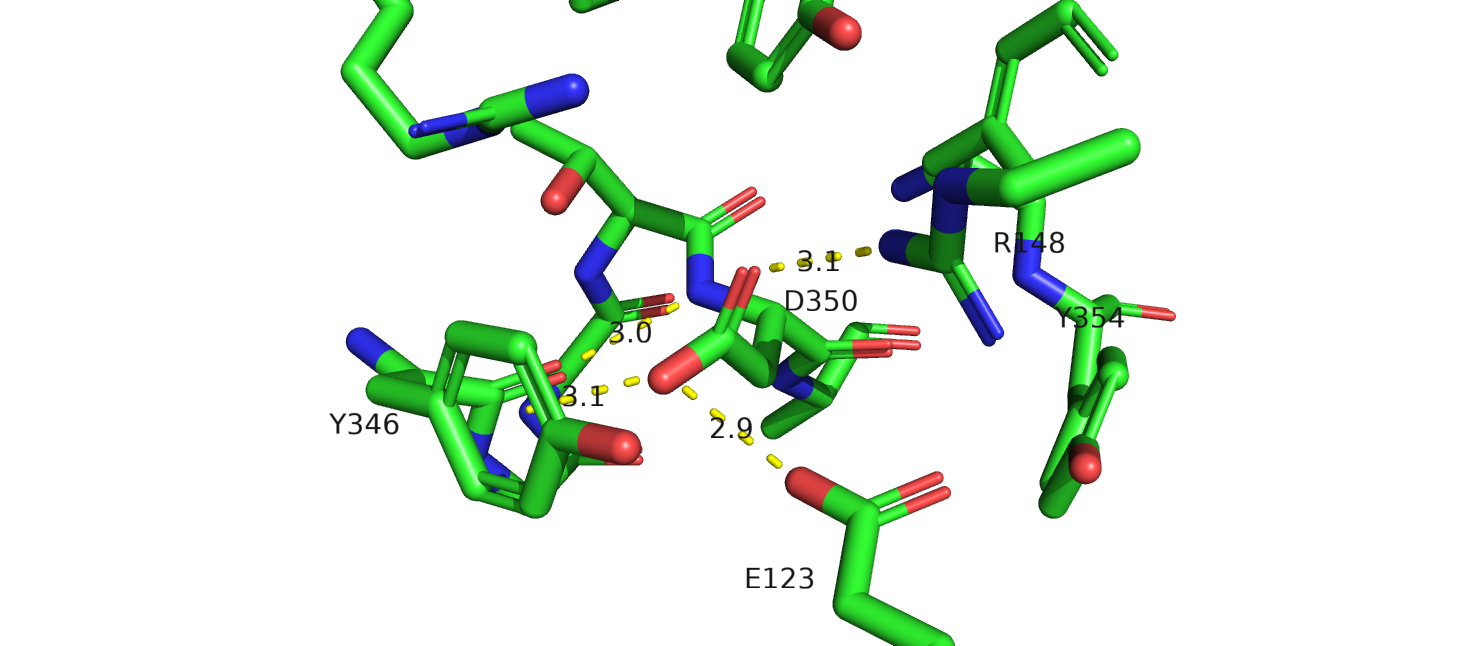
В структуре 6os9 аспарагин также взаимодействует с аргинином-148 (не показан на рис.19б), кольцом тирозина, глутаматом. Отрицательный заряд
мутации приведет к потере пи-водородной связи. Это может привести к тому, что тирозин повернется в сторону лиганда. Однако как именно это повлияет
на связывания лиганда, я не могу сказать. Этот тирозин достаточно далеко расположен от лиганда.
Так что мне кажется, что мутация N344D не повлияет на функцию белка (нейтральная).
Рис.19 c. 6os9, остаток аспарагина-344(350 в PDB), мутированный на аспартат, тирозин и лиганд
Структурный поиск
Я также решила провести структурный поиск с помощью PDBeFold. Поиск я производила по R цепи структруры 6os9 (рецепторная часть). Было найдено 97 хитов.
Эти хиты до 13 все комплексы с рецептором нейротензина 1, а потом начинаются другие рецепторы. Я сделала выравнивание первых 9 хитов.
Из минусов PDBeFold - когда я хотела скачать файл pdb с новыми координатами, открывалась просто страница с файлом котоую нельзя было скачать,
приходилось копировать и вставлять в новый файл, это мне не понравилось, поэтому я скачала только первые 4 структры. Еще минус - не работала визуализация (рис. 20).
1 хит - это уже описанная выше 6os9, второй - 6osa, тоже человеческий рецептор, а 3 и 4 - 7l0s и 7l0r, очень похожие структуры, рецепторная часть у них крысиная (на рис. 21 розовый и желтый, соответственно).
Рис.20 Немного из выравнивания PDBeFold
Рис.21. Выравнивание первых 4 хитов PDBeFold.
5 хит, кстати, был в выдаче BLAST, но они примерно одинаковые эти все 7L0* (из одной статьи). Интересно, кстати,
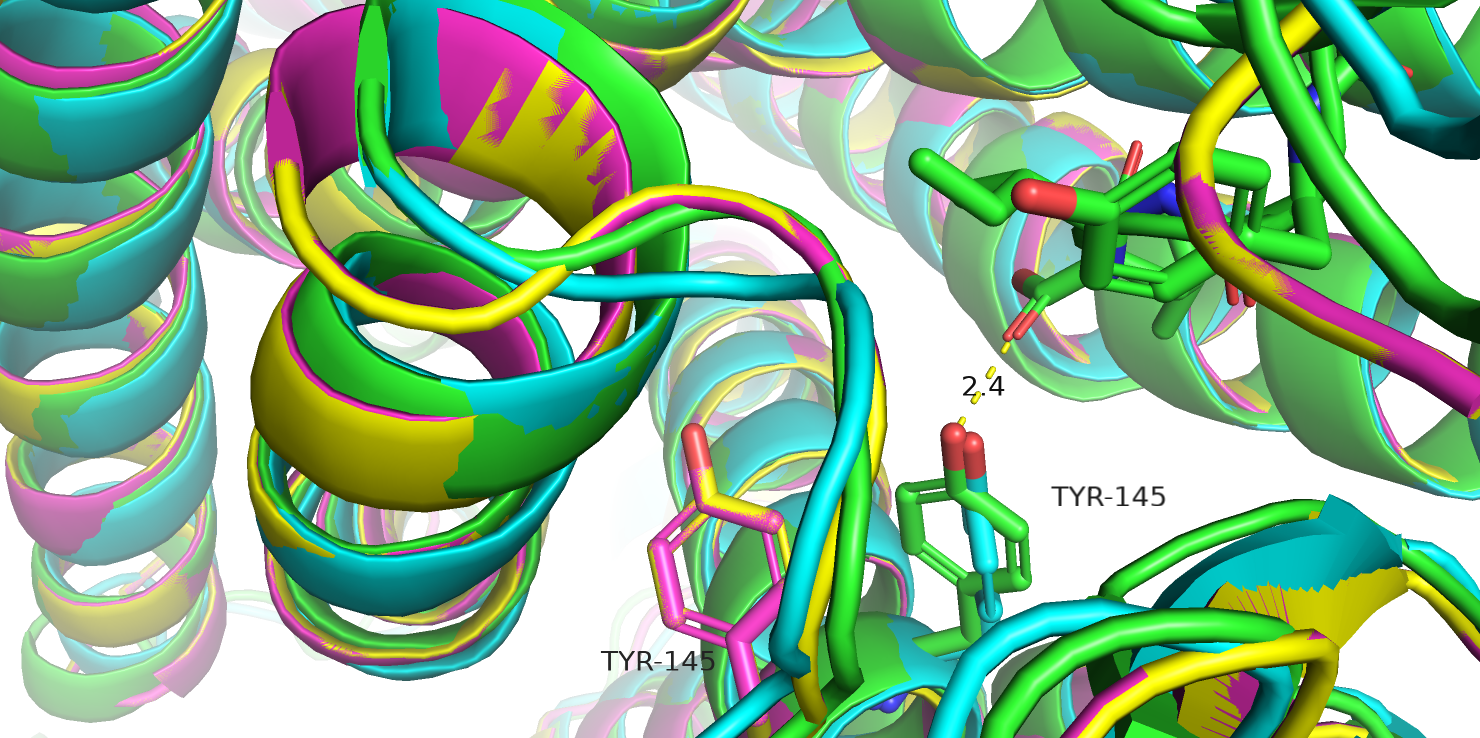
на рис. 22 можно заметить, что крысиный 145 тирозин, мутацию в котором я ранее уже рассматривала, находятся немного дальше от лиганда,
и врядли могут с ним взаимодействовать. В этих структурах, конечно, внесены 10 замен, но я посмотрела их, они вообще не находятся в непосредственной
близости от этого остатка, и как-то напрямую на его положение вряд ли влияют. Так что, учитывая родство крыс и грызунов, из которых взят
исследуемый белок, можно предположить, что в исходном белке этот 145 тирозин тоже не взаимодействует с лигандом, а это уже может
повлиять на результат мутации в этом остатке, т.е. мутация может оказаться и нейтральной.
Я посмотрела на последовательность исходного белка, крысиных и человеческих, и оказалось, что в исходном белке там есть такое GYYF,
где второй Y - это как раз 145. Нумерация этих остаток отличается у крыс и у человека на 1 позицию.
Посмотрим на рис. 23-26. Второй
тирозин от глицина в этом месте скорее всего взаимодействует с лигандом, а мутация, которая ведет к потере этого взаимодействия,
ведет и к ухудшению связывания и потери функции (может частичной). Правда, тут надо отметить, что на рисунках лиганд показан из структуры
6os9, но скорее всего, лиганд так и располагается в крысах. Я посмотрела для структуры 7l0s, там тоже есть взаимодействие, там даже как-то
слишком близко тирозин и лиганд расположены (рис. 27 - рядом с 22)
Рис.22. Тирозин-145 у крыс (роз., желт.) и у человека (гол., зел.)
Рис.27. Тирозины-145-146 и лиганд у крысы из 7l0s
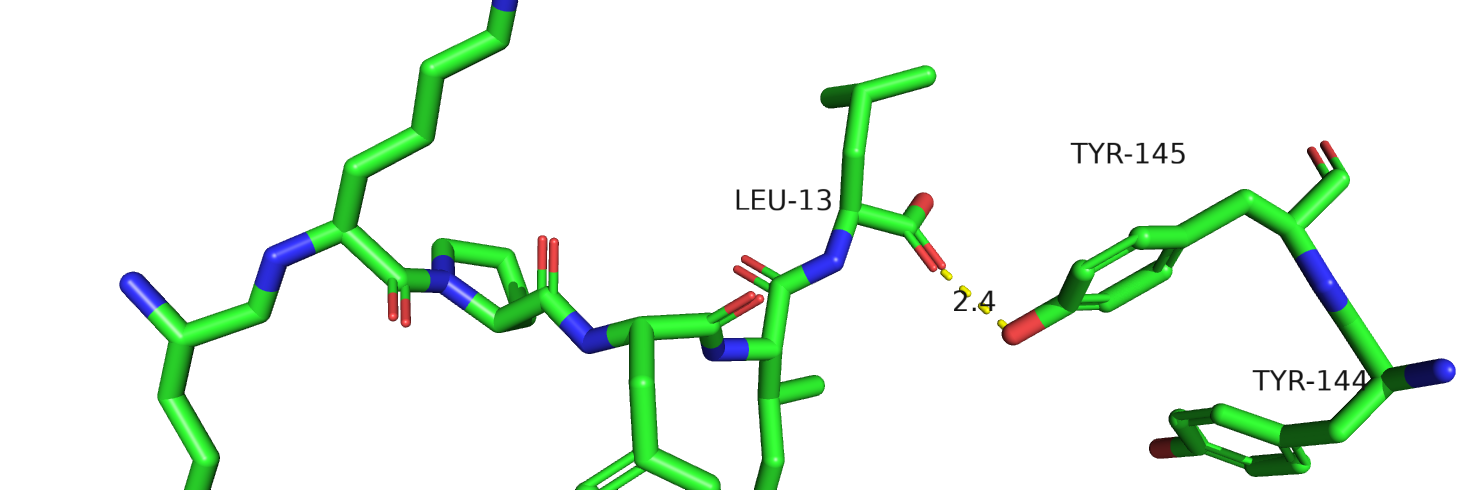
Рис.23. Тирозины-144-145 у человека из 6os9
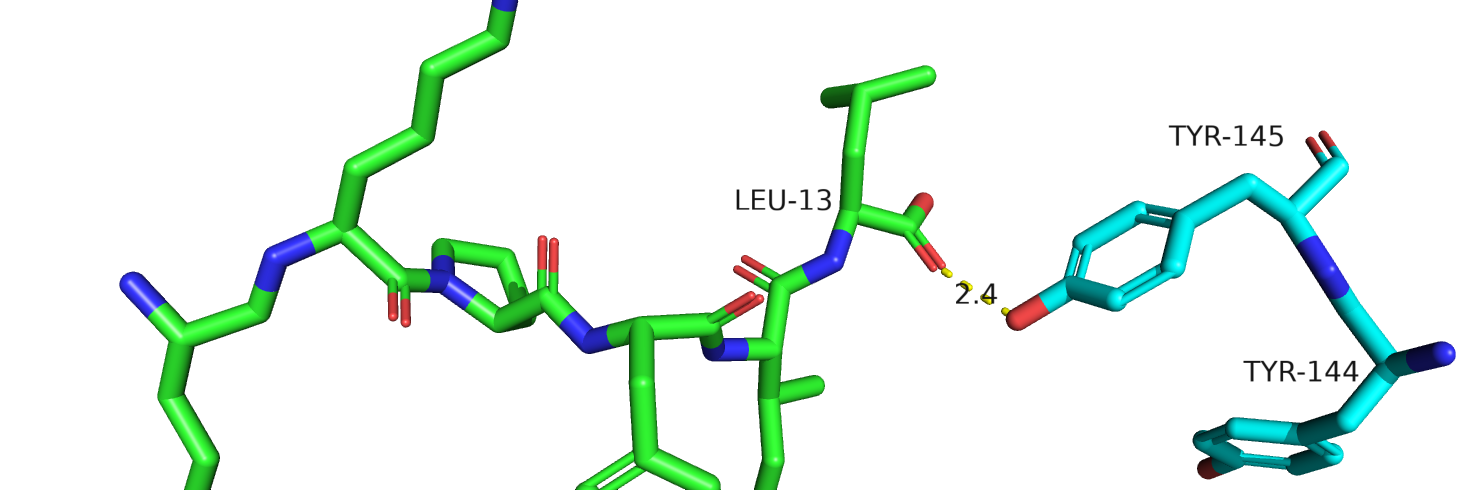
Рис.24. Тирозины-144-145 у человека из 6osa
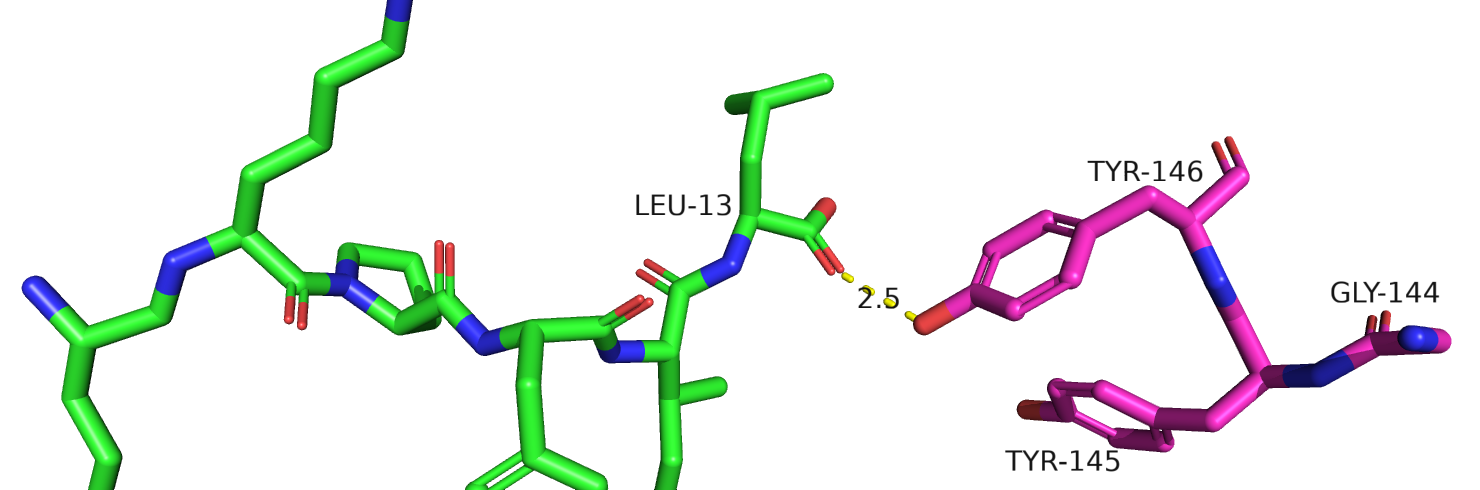
Рис.25. Тирозины-145-146 у крыс из 7l0s
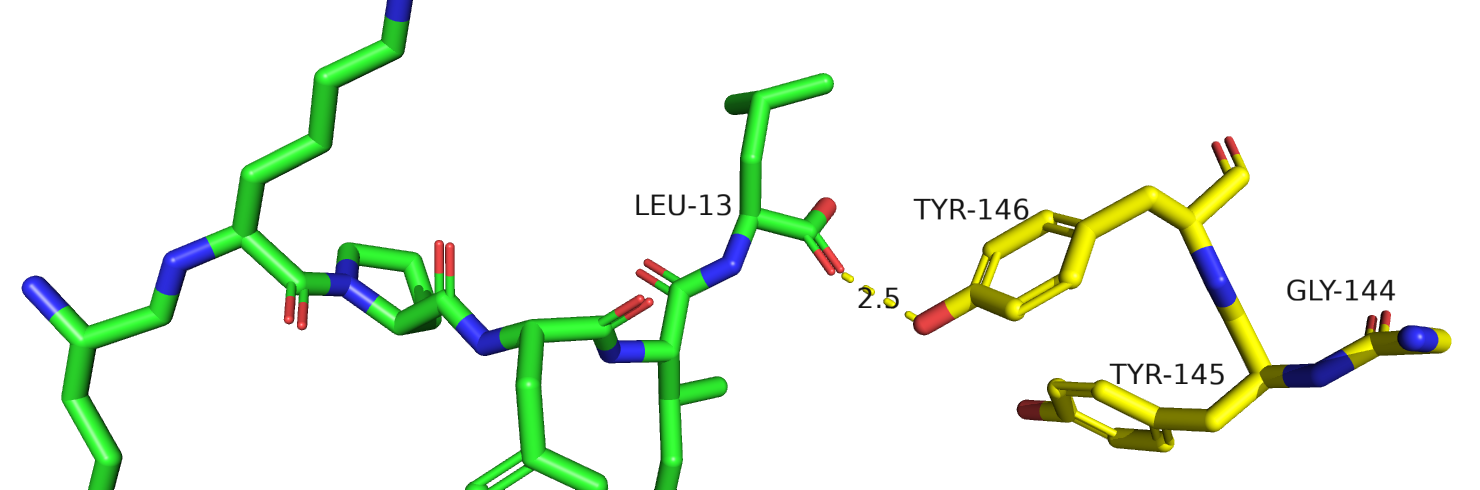
Рис.26. Тирозины-145-146 у крыс из 7l0r
Вывод
Я надеюсь, я смогла ответить на вопрос о том, как повлият мутации на функционирование белка. По моим расчетам,
мутация Y145F будет негативной, так как приводит к ухудшению связвания лиганда, а мутация R317H приведет также к негатвным последствиям, как из-за
того, что потеряно возможное взаимодействие, так и потому, что мутантный остаток может вызвать конформационные изменения в сайте связывания лиганда.
Мутация N344D, по моему мнению, является нейтральной, так как не влияет на свзявание лиганда и не вызывает изменения в структуре.
Вообще, хотелось бы добавить, что, так как все 3 мутации находились в кармане связывания нейротензина, было бы интересно еще посмотреть и на мутации
остатков, участвующих в связывании с другими белками комплекса, ведь смысл связывания лиганда рецептором заключается в том, чтобы передать
сигнал дальше на другие белки каскада.