В этом задании мне достался белок, формирующий канал в плазматической мембране клеток и внешней мембране митохондрий.
Несмотря на сложности кристаллизации мембранных белков, для него была получена РСА структура 6G6U с разрешением 2.74 ангстрем,
для сравнения была взята структура 6TIQ, полученная методом ЯМР и представленная 20 моделями.
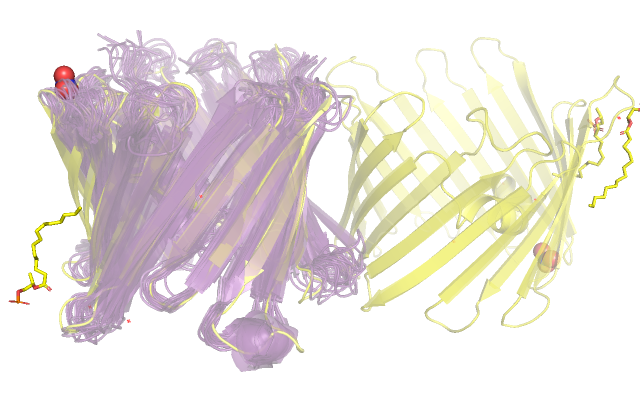
Рис.1 Общий вид структур, полученных методами РСА (желтым цветом) и ЯМР (фиолетовым цветом).
На рис.1 мы видим, что для РСА структуры присутствуют молекулы воды и растворителя, чего не наблюдается в структуре ЯМР,
так как там зачастую применяют дейтериевую воду, не дающую сигнала. Для структуры, полученной методом рентгено-структурного анализа
наблюдаются две молекулы кристаллической ячейки, в то время как для ЯМР структуры мы видим набор моделей, соответствующих
ограничениям на расстояния, полученным экспериментально и взятым из литературы. В плане сходства структур можно заметить, что
бета-листы, составляющие бочонок, довольно хорошо совпадают, хуже это наблюдается для альфа-спирали в центре и совсем плохо
для петлей между бета-листами, что может объясняться увеличением степеней свободы, а следовательно, подвижности для таких остатков,
что приводит к размытию электронной плотности в случае РСА эксперимента и появлению большого числа равных по энергии положений в случае
ЯМР.
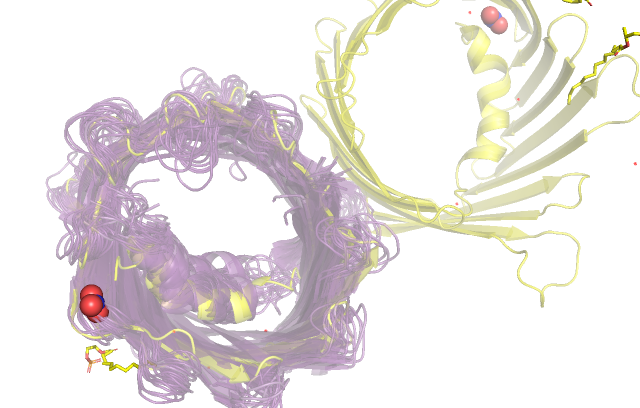
При ближайшем рассмотрении сразу становятся видны новые различия. Так, в ЯМР структуре присутствуют водороды в силу проведения эксперимента (их
и детектируем), в структуре РСА при таком разрешении они не наблюдаются и могут быть очень грубо восстановлены визуализатором. Также в случае
рентгено-структурного анализа несколько остатков на концах белка не были разрешены, вероятно, в силу их большой подвижности.
В структуре бета-листов на их концах обнаружено много заряженных и полярных аминокислот, часть из которых обращены внутрь бочонка для переноса
ионов через мембрану и не образуют водородных связей или солевых мостиков в обеих структурах, в то время как часть должны, казалось бы,
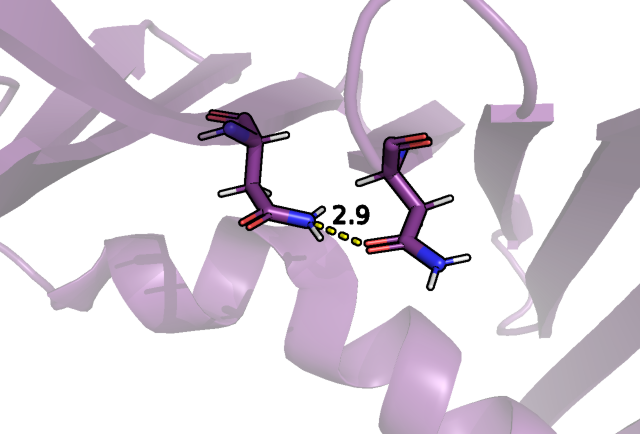
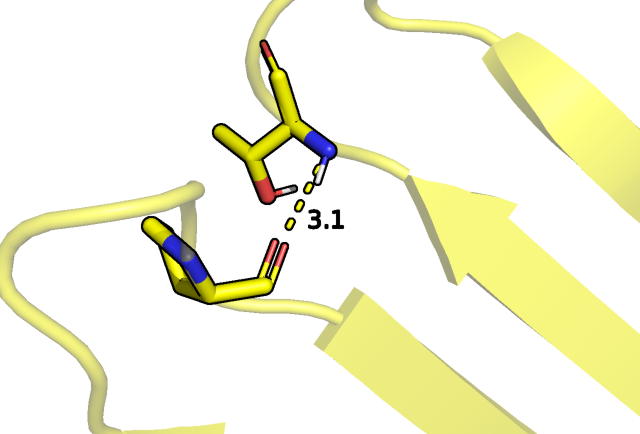
участвовать в образовании упомянутых связей, так, в случае ЯМР структуры это было показано для остатков ASN 168 и ASN 185 в части структур:
Рис.2 Наличие/отсутсвие взаимодействий между близко расположенными полярными остатками.
Водородная связь между остаками аспарагина 168 и аспарагина 185 показана желтым пунктиром.
Такое различие может быть связано с плохим вписыванием остатков аспарагина в электронную плонтость в РСА эксперименте в силу разрешения и того, что трудно
различить кислород и азот боковой группы этого остатка между собой в контексте ЭП (инверсия ASN).
Задание 2. Prody и RMSF
В этом задании предлагалось изучить параметр RMSF (Root mean square fluctuation), показывающий стандартное отклонение
координат атомов по времени, в данном случае в качестве динамики мы берем ансамбль структур ЯМР, предполагая, что он отражает подвижность
белка в растворе:
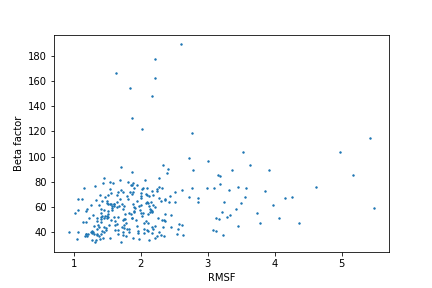
Рис.3 Зависимость среднего RMSF по остатку от его среднего В-фактора.
На графике на рис.3 можно заметить наличие определенного тренда: чем больше RMSF, тем больше В-фактор, однако на начальном отрезке
RMSF наблдаются и большие значения структурного фактора, что может говорить о том, что эти значения характеризуют не динамическую подвижность
РСА структуры, а размытие ее ЭП, что подтверждается показателями качества модели РСА. Так, разрешение 2.7 А можно считать удовлетворительным,
значение Rfree равняется 0.287 > 0.2, что не является хорошим показателем.
Задание 3. Водородные связи
В этом задании надо было рассмотреть представленность различных водородных связей в моделях. Для начала возьмем водородные связи остова в
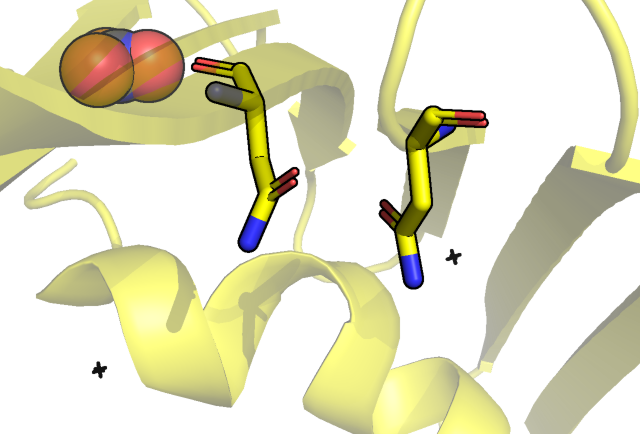
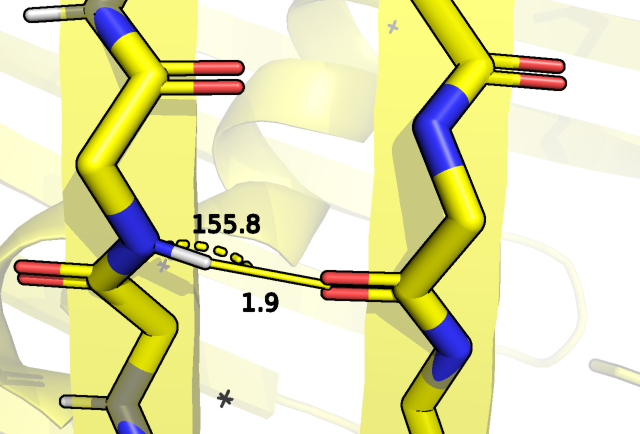
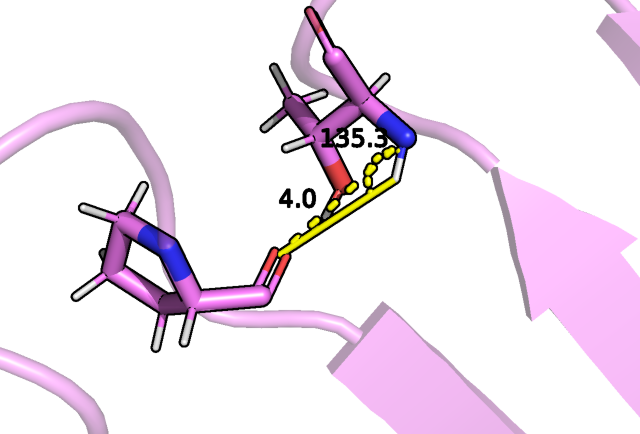
бета-листах бочонка, например, между кислородом изолейцина 85 и азотом фенилаланина 99:
Рис.4 Водородная связь между остовом ILE 85 и PHE 99.
Как мы видим в случае ЯМР структуры, наблюдаемая в РСА водородная связь не для всех моделей имеет место быть (брался критерий угол больше 150 градусов,
расстояние водород акцептор 2 ангстрема плюс минус 0.5 ангстрем). В этом случае смотрели на расстояние водород акцептор, что в рамках остова
допустимо, так как геометрия водорода задается однозначно.
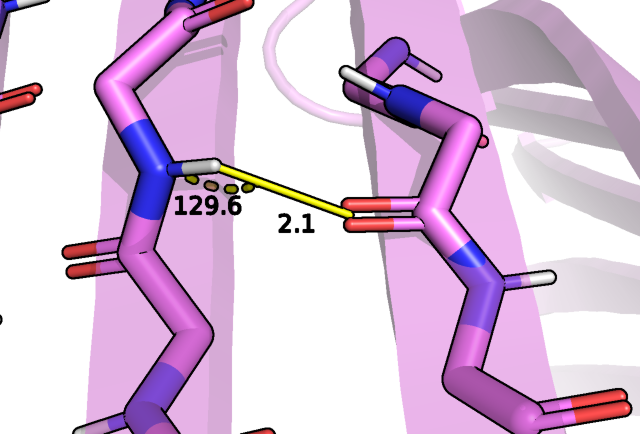
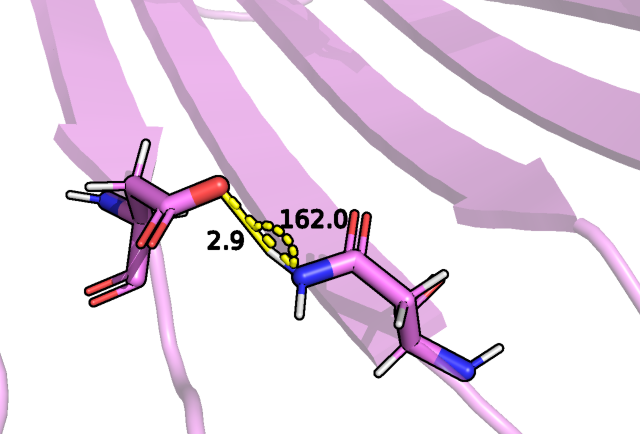
Далее была рассмотрена водородная связь для боковых групп бета-листов, например, между азотом аспарагина 111 и кислородом
аспартата 130:
Рис.5 Водородная связь между боковой цепью ASN 111 и ASP 130.
Здесь на рисунке 5 мы видим только одну модель из набора ЯМР структур, которая может иметь водородную связь.
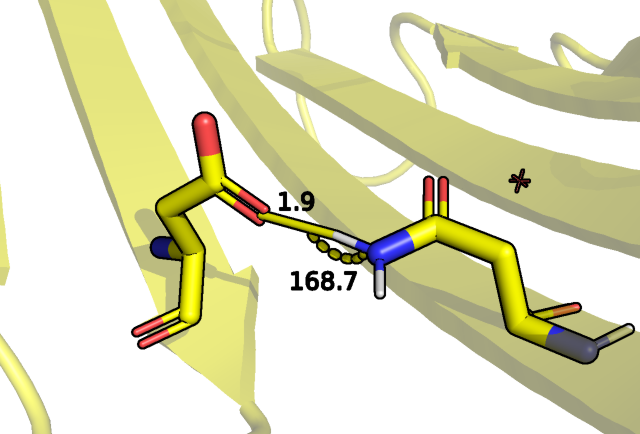
Наконец были взяты остатки, входящие в петлю структуры, а именно, остовный азот треонина 159 и остовный кислород пролина 136:
Рис.6 Водородная связь между остовом THR 159 и PRO 136.
Как мы видим на рисунке выше здесь тоже не для всех ЯМР структур было показано возможное наличие этой связи (критерий брался, как угол больше 150 градусов,
расстояние акцептор донор 3 ангстрем плюс минус 0.5 ангстрем).
Таблица 1. Расстояния между донором и акцептором водородной связи (для первого варианта между акцептором и водородом).
Номер связи
Расстояние для РСА модели (А)
Процент моделей ЯМР со связью (%)
Минимальное расстояние в ЯМР (А)
Максимальное расстояние в ЯМР (А)
Медианное значение в ЯМР (А)
1
1.9
85
1.7
2.1
1.8
2
2.9
5
2.7
9.6
6.35
3
3.1
5
3.3
5.1
3.9
Из таблицы 1 можно сделать вывод, что в случае остатков, находящихся в стабильных структурах таких, как бета-листы, в случае остова ЯМР и РСА дают
похожие результаты, что согласуется с теорией (взаимодействия образующие бета-лист должны быть очень стабильными). Для боковых цепей тех же остатков, однако,
в данном случае результаты ЯМР сильно разошлись с РСА, что, вероятно, можно объяснить неправильным вписыванием остатков в ЭП авторами модели.
Для остатков петли наблюдается то же самое, но здесь это скорее всего объясняется большой подвижностью этой части структуры, так как атомы остова
вписать неправильно представляется менее вероятным.