ФББ 2013-2014


ФББ 2013-2014
Предсказание парных выравниваний.
Проект Jalview с выравниваниями и разметкой можно скачать здесь.
Парное выравнивание - это выравнивание двух аминокислотных последовательностей друг относительно друга. Оно строится с той же целью, что и множественное выравнивание - то есть по парному выравниванию пытаются оценить гомологию данных последовательностей. Однако работа с парным выравниванием всегда предполагает ориентацию на множественное выравнивание белков этого семейства, это нужно для более точной оценки консервативности.
Для выполнения заданий я выбрала это выравнивание и совмещение из предложенных. Само выравнивание можно увидеть здесь. Затем я открыла это выравнивание в редакторе JalView, общий вид проекта можно увидеть на рисунке 1.
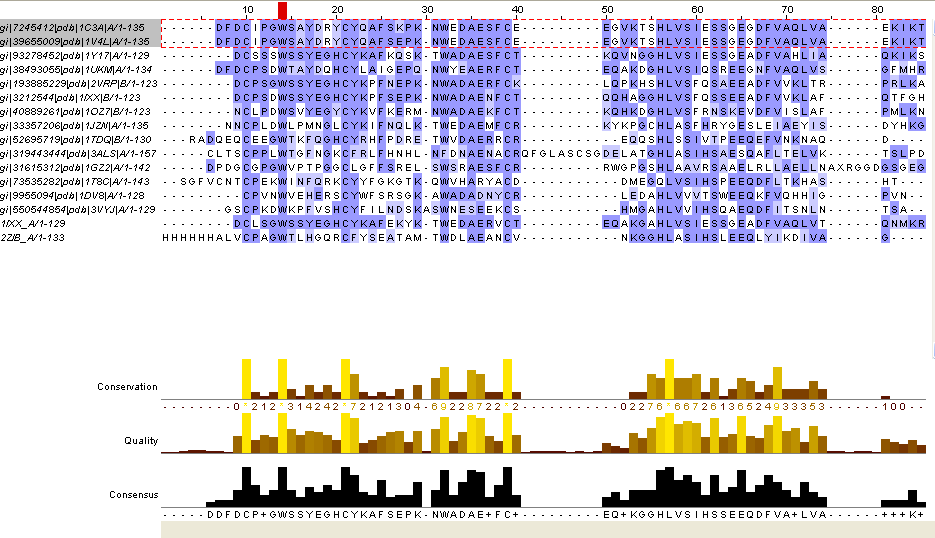
Рис.1. Множественное выравнивание 16-ти последовательностей.
Для того, чтобы более наглядно увидеть родственные связи между белками, я построила дерево множественного выравнивания (рисунок 2). По нему видно, что последовательности, которые я буду выравнивать, не очень гомологичны друг другу (по крайней мере по сравнению с остальными). 1IXX - это белок из яда змеи хабу, который связывается с факторами свёртывания крови IX/X. 2ZIB - это Ca-независимый белок-антифриз, выделенный из Brachyopsis rostratus (это костистая рыба).
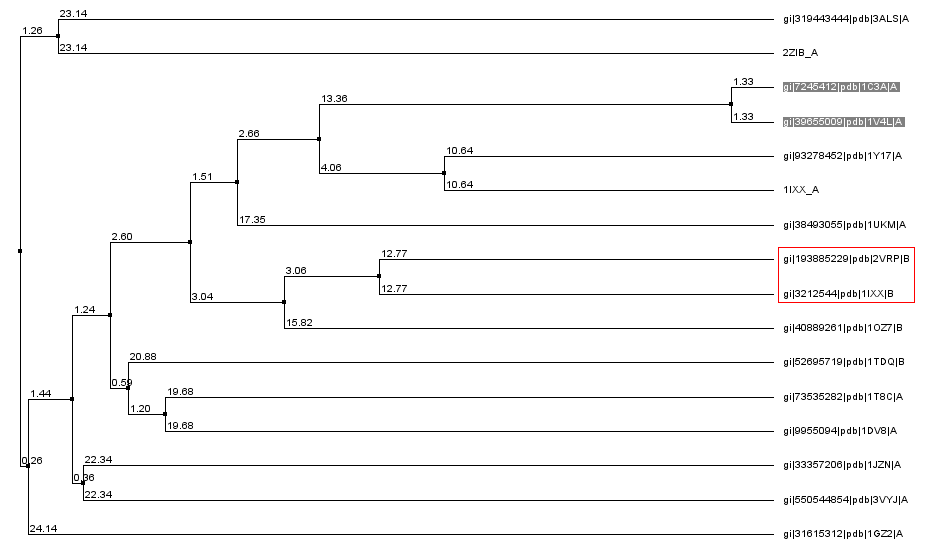
Рис.2. Дерево множественного выравнивания. Выбранные для парного выравнивания последовательности обведены в красный прямоугольник.
По заданию 1 необходимо построить вручную выравнивание двух последних последовательностей из множественного выравнивания относительно друг друга. Результат:
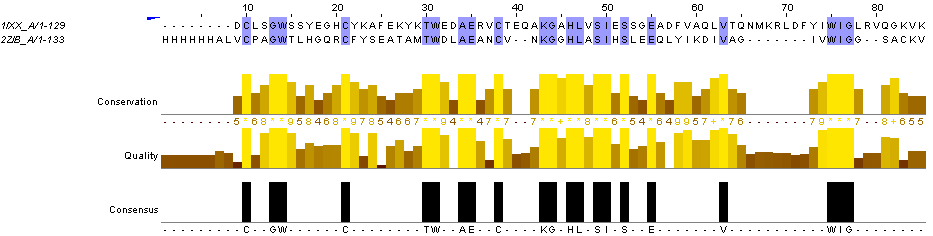
Рис.3.Парное выравнивание последовательностей 1IXX_A и 2ZIB_A, построенное с помощью средств Jalview.
Для выполнения задания 2 я воспользовалась командой needle на kodomo. Программа needle использует алгоритм Нидлмана-Вунша для построения глобального выравнивания двух последовательностей. Глобальное выравнивание подразумевает гомологию последовательностей по всей длине, туда включаются обе последовательности целиком. Сначала я получила парное выравнивание двух последних последовательностей в формате по умолчанию, его можно скачать здесь. Затем, дописав опцию -aformat3 fasta я получила output файл в формате fasta, с ним можно ознакомиться здесь. Выравнивание в формате fasta я визуализировала в JalView, раскраска BLOSUM62 (рисунок 4).
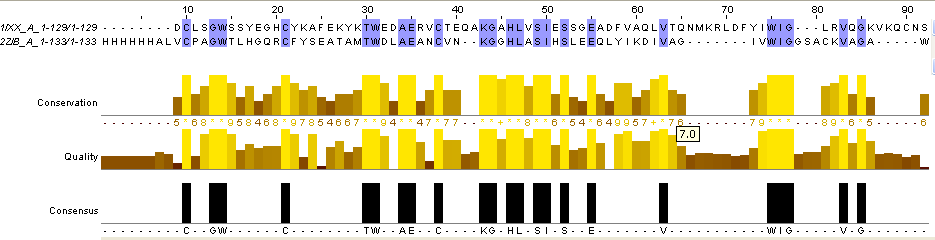
Рис.4.Парное выравнивание двух последних последовательностей (1IXX_A и 2ZIB_A) из множественного выравнивания, построенное с помощью программы needle на kodomo. Раскраска BLOSUM62
Далее я построила наилучшее локальное выравнивание двух последних последовательностей (1IXX_A и 2ZIB_B) в формате fasta, используя для этого команду water на kodomo. Локальное выравнивание строится, когда в последовательностях есть негомологичные участки, тогда они исключаются и выравнивание идёт между гомологичными участками. Для получения локального выравнивания используют алгоритм Смита-Ватермана. Результат в формате fastа можно увидеть здесь. В редакторе Jalview это выравнивание выглядит так:
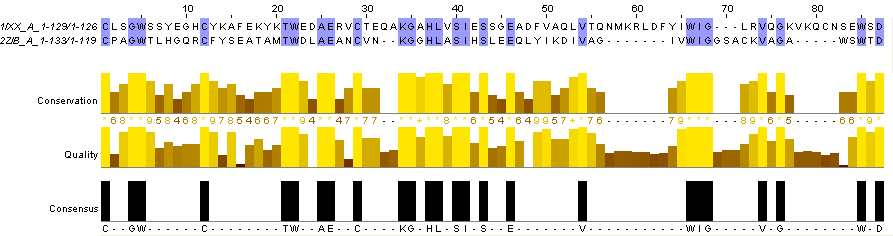
Рис.5.Наилучшее локальное выравнивание двух последовательностей (1IXX_A и 2ZIB_B), открытое в редакторе Jalview. Раскраска BLOSUM62.
Для выполнения задания 4 я взяла 2 последовательности: 1IXX_A (с ней же велась работа в заданиях 2 и 3) и 3GL3_A (с этой последовательностью я работала в более ранних практикумах). Последовательности заведомо негомологичны: 1IXX_A - это змеиный яд, а 3GL3_A - это прокариотный белок дисульфидного обмена. Полученные результаты глобального и локального выравниваний:
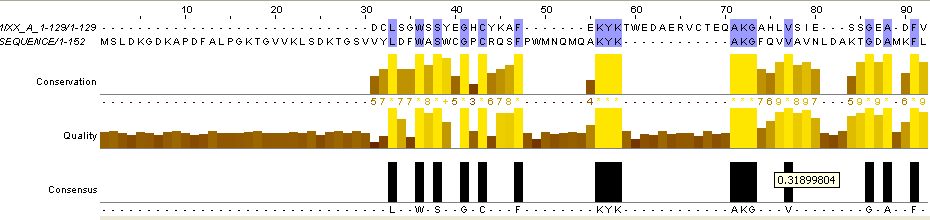
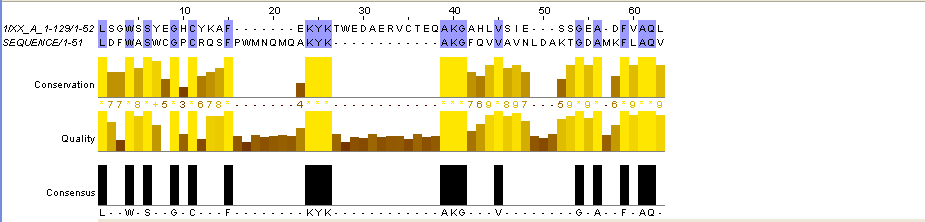
Рис.6. Выравнивание заведомо негомологичных последовательностей белков с pdb идентификаторами 3GL3 и 1IXX. На верхнем рисунке - глобальное выравнивание, на нижнем - локальное.
Сравнение параметров выравниваний приведено в таблице 1. Как видно, выравнивание вручную даёт менее точные результаты, чем глобальное и локальное выравнивание с помощью алгоритмов. Сравнивать же локальное и глобальное выравнивание нельзя, но стоит отметить, что в случае гомологичных последовательностей длина глобального и локального выравниваний отличаются не намного, в то время как для заведомо негомологичных последовательностей длина локального выравнивания совсем маленькая. В выравниваниях заведомо негомологичных последовательностей наблюдается заметно больше гэпов, с помощью них программа пыталась найти хоть какое-нибудь сходство. Число и процент совпадений в глобальных выравниваниях гомологичных и негомологичных последовательностей различаются не сильно за счёт того, что программа вставила много гэпов в выравнивание негомологичных последовательностей. В локальном выравнивании негомологичных последовательностей мы видим самое большой процент совпавших остатков, однако из этого нельзя сделать вывод о гомологии, т.к. это сходство достигается за счёт большого процента гэпов и оно наблюдается на коротком участке по сравнению с глобальным выравниванием.
Табл.1 Параметры проведённых парных выравниваний.
|
Длина выравнивания (число колонок) |
Число совпадений |
Процент совпадений |
Число сходных остатков |
Процент сходных остатков |
Число гэпов |
Процент гэпов |
Число открытий гэпов |
|
|
Выравнивание вручную в Jalview (рис. 3б) |
144 |
31 |
21,5% |
45 |
32,25% |
26 |
18% |
5 |
|
Глобальное выравнивание с помощью needle (рис.4) |
148 |
35 |
23,6% |
56 |
37,8% |
34 |
23% |
6 |
|
Локальное выравнивание c помощью water (рис.5) |
134 |
35 |
26,1% |
56 |
41,8% |
23 |
17,2% |
5 |
|
Глобальное выравнивание 2х заведомо негомологичных белков (рис.6) |
190 |
29 |
15,3% |
48 |
25,3% |
99 |
52,1% |
7 |
|
Локальное выравнивание 2х заведомо негомологичных белков (рис.6) |
63 |
18 |
28,6% |
29 |
46% |
23 |
36,5% |
3 |
Выполняя задание 6, я добавила глобальное выравнивание, полученное с помощью needle, к выравниванию, которое я получила вручную. Сразу обнаружилось высокое сходство на начальном участке выравниваний, там не пришлось ничего двигать (рисунок 7).
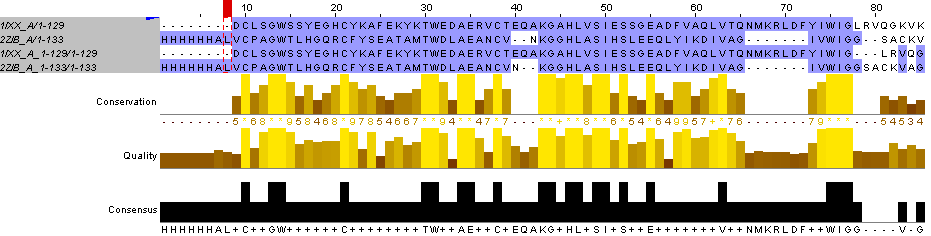
Рис.7. Высокое сходство двух парных выравниваний одних и тех же последовательностей на начальном участке.
Однако на концевом участке выравнивания совмещались не так хорошо. Чтобы получить совмещение я вставила 3 гэпа в обе последовательности первого выравнивания так, чтобы это не повлияло на само выравнивание. Получилось хорошее совмещение (до - рисунок 8, после - рисунок 9).

Рис.8. Концевой участок выравниваний, на котором автоматическое выравнивание сильно отличается от полученного вручную.
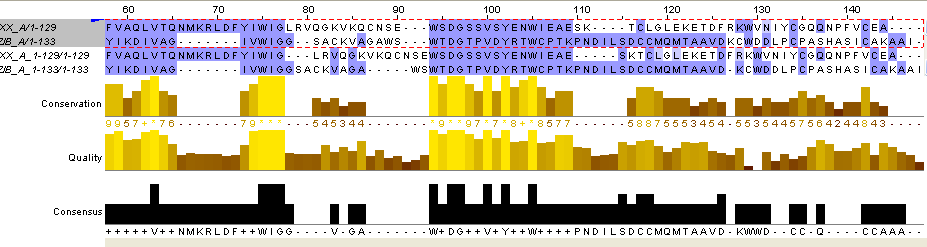
Рис.9. Совмещение двух выравниваний на концевых участках путём добавления гэпов в первое выравнивание.
Проверять правильность выравнивания я буду с помощью программы SupCheck, которая совмещает пространственные структуры двух белков. Это может помочь найти ошибки в выравнивании (обычно они есть) и установить гомологию между участками белка. Окно Rasmol с совмешением двух структур показано на рисунке 10. Выравнивание, полученное с помощью needle, в целом более соответствует пространственному совмещению, чем выравнивание с помощью удаления гэпов.

Рис.10. Совмещение двух пространственных структур белков 1IXX_A и 2ZIB_A в Rasmol.
Для каждого выравнивания я отдельно посмотрела его соответствие с совмещением. Количество ошибок в выравнивании вручную: 40 ошибок I рода (остатки, расположенные в одной колонке выравнивания, на структуре не совмещены), 4 ошибки II рода (хорошо совмещающиеся в пространстве остатки не расположены в одной колонке выравнивания). Количество ошибок при выравнивании needle: 26 ошибок I рода, 3 ошибки II рода. На рисунке 11 показан участок выравнивания с ошибками I рода. На самом деле, обе цепи образуют сходные в пространстве структуры на участке, выделенным красным в Jalview, но эти структуры расположены под разным углом, поэтому формально не совмещаются. Однако и на этом участке можно говорить о гомологии из-за двух причин: совпадают довольно редкие аминокислоты и при наложении независимо от других частей белков эти участки совпали бы.
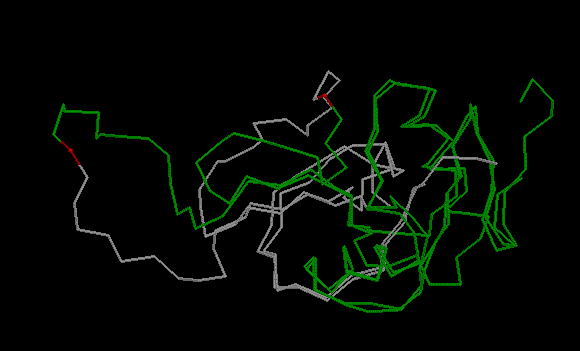
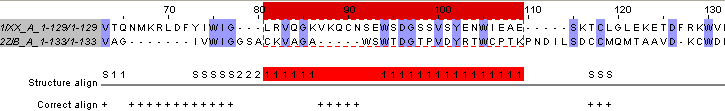
Рис.11. Совмещение двух пространственных структур белков 1IXX_A и 2ZIB_A в Rasmol.