2013-2014


2013-2014
База данных KEGG
KEGG PATHWAY - сегмент базы данных KEGG, в котором хранится информация о метаболических путях клетки. Информация организована в виде графов, в узлах которых находятся химические вещества (продукты реакций), а на рёбрах указано направление реакции и фермент, её катализирующий. Для работы с этой базой был выбран Цикл трикарбоновых кислот (цикл Кребса). Он изображён на рисунке 1.
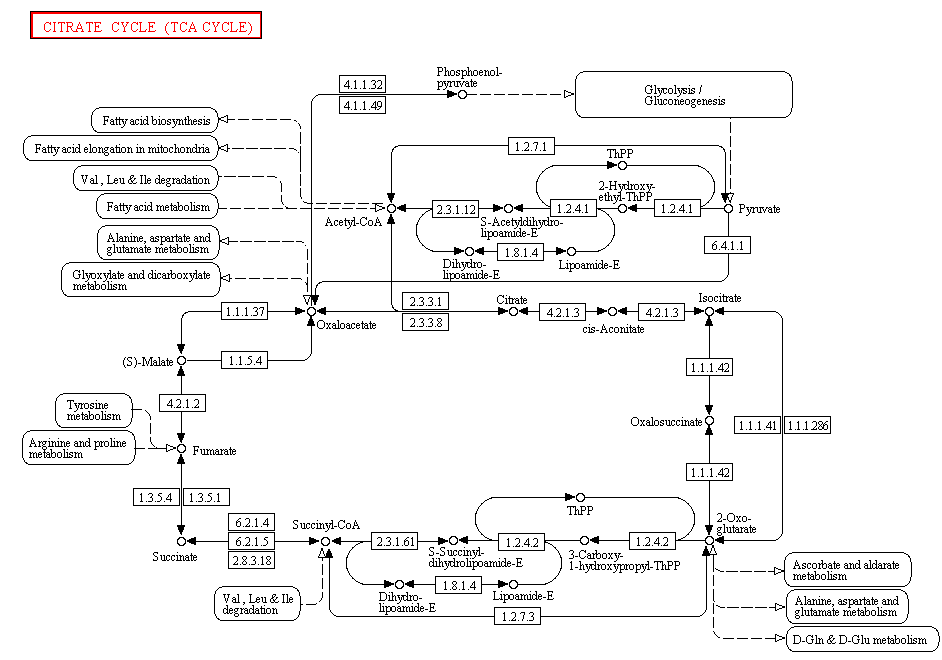
Рис.1. Цикл трикарбоновых кислот в базе данных KEGG. Он же на сайте базы - ссылка.
Цикл Кребса является центральным путём окисления соединений углерода. Он представляет из себя 8 последовательных реакций, в первую реакцию вступает лимонная кислота и она же образуется в результате восьмой реакции, замыкая цикл. Лимонная кислота, в свою очередь, получается из оксалоацетата и ацетил-КоА - конечного продукта второй стадии катаболизма углеводов, жиров и аминокислот. Лимонная кислота содержит 6 атомов углерода, в цикле она несколько раз подвергается дегидрированию и 2 раза - декарбоксилированию, после этих превращений из неё регенерируется оксалоацетат. Как видно из рисунка 1, в цикл Кребса также формально включают образование самого ацетил-КоА из пирувата в ходе реакции, катализируемой пируватдегидрогеназой. Цикл Кребса протекает в матриксе митохондрий. Его конечными продуктами являются углекислый газ и высокоэнергетические электроны в форме NADH, которые потом поступают на цепь переноса электронов во внутренней мембране митохондрий.
Цикл Кребса характерен для всех аэробных организмов. Молекулярный кислород в самом цикле не используется, однако он является единственным эффективным способом снятия электронов с NADH с образованием NAD+ (NAD+ необходим для цикла). Другими продуктами этого метаболического пути являются FADH2 и GTP.
Кратко о реакциях цикла Кребса:
С помощью KEGG PATHWAY можно понять, с какими другими метаболическими путями связан цикл Кребса:
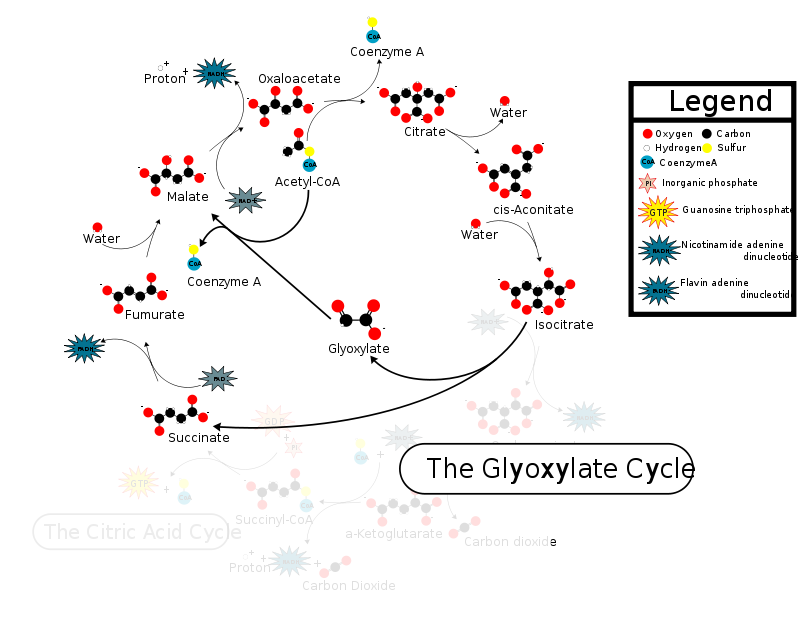
Рис.2 Модификация цикла Кребса - глиоксилатный путь.
Глиоксилатный путь известен для всех, кроме животных (хотя у них, предположительно, он тоже есть).Для того, чтобы оценить распространённость цикла Кребса среди живых организмов было выбрано 2 эукариота (Ornithorhynchus anatinus - утконос, Giardia lamblia), 3 бактерии (Chlorobium tepidum, Lactobacillus sp., Photorhabdus luminescens), 1 архея (Methanocladococcus jannaschii). Наличие ферментов цикла Кребса у данных организмов показано на рисунке 3 (картинки меняются по клику). Если фермент присутствует, то он выделен зелёным.
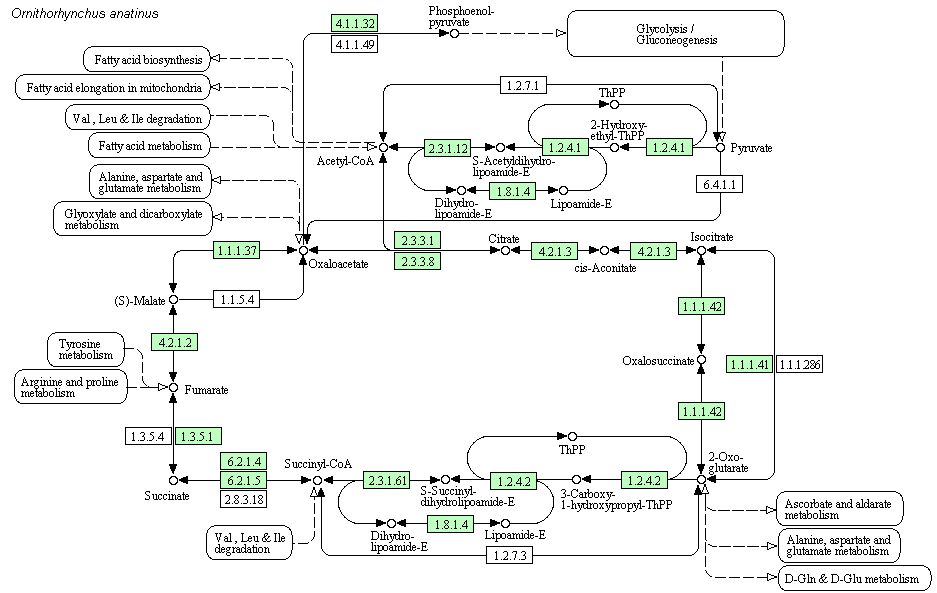
Рис.3. Ферменты цикла Кребса у выбранных организмов. Зелёным отмечено наличие ферментов. Картинки меняются по клику.
Как видно из рисунка 3, полный цикл Кребса может реализоваться у утконоса (Ornithorhynchus anatinus) и у Photorhabdus luminescens, так как у них есть
нужный фермент для каждой стадии реакции. У Chlorobium tepidum присутствует достаточно много ферментов цикла Кребса, однако отсутствуют
ферменты для синтеза сукцинил-КоА. Скорее всего, этот организм использует реакции и ферменты цикла Кребса в синтетических целях, а не в
целях получения энергии. У
Получившийся результат был вполне предсказуем: у аэробных организмов работает стандартный цикл Кребса, а у анаэробных - он либо совсем не работает, либо его реакции используются в других процессах.
KEGG Reaction
Каждая реакция данного метаболического пути содержится в KEGG под отдельным идентификатором. На странице реакции можно получить исчерпывающую информацию о ней: какие катализаторы используются, в каких метаболических путях участвует, кем и когда открыта. Помимо конечного и начального продукта реакции также подписываются кофакторы, необходимые для протекания процесса. Для выполнения задания я выбрала реакцию декарбоксилирования изоцитрата, для которой необходим NAD+. Реакцию в базе данных можно найти под номером R00709.
На рисунке 4 показано представление этой реакции в базе данных KEGG. В ходе реакции происходит не только декарбоксилирование субстрата, но и восстановление NAD+ до NADH. Уравнение реакции: Isocitrate + NAD+ <=> 2-Oxoglutarate + CO2 + NADH + H+.
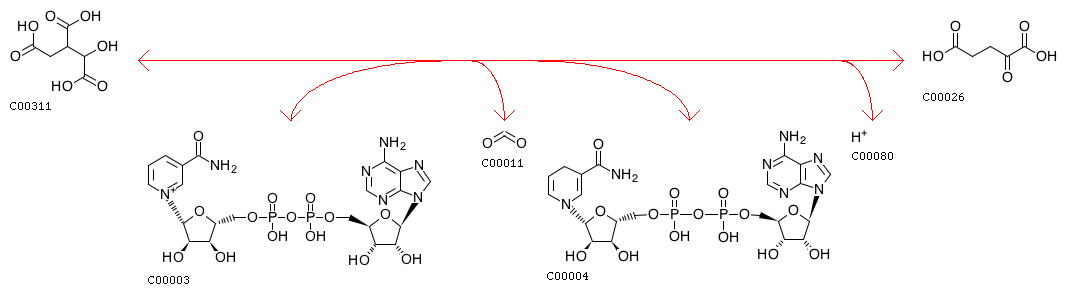
Рис.4 Схема реакции декарбоксилирования изоцитрата до альфа-кетоглутарата.
Место выбранной реакции на общем метаболическом пути показано на рисунке 5.
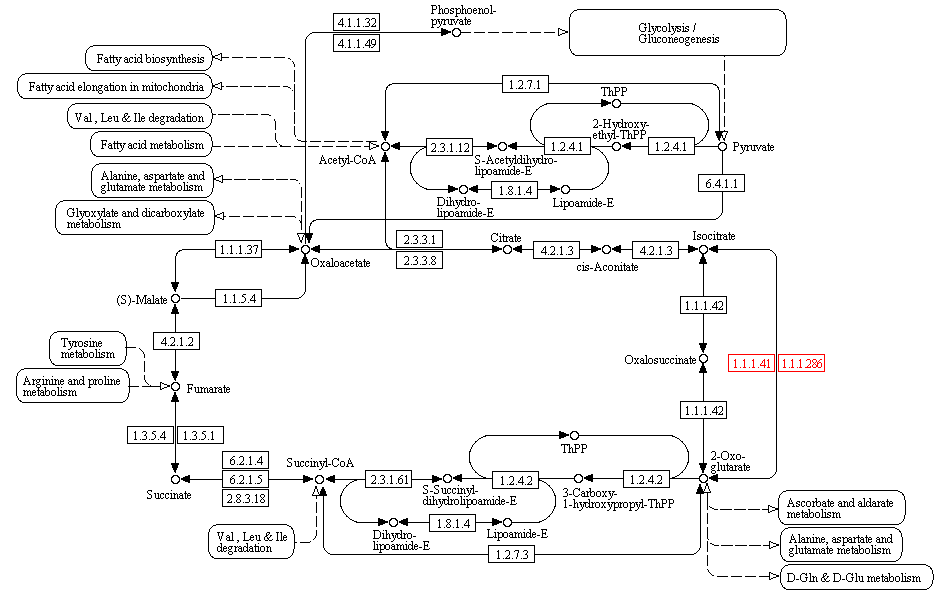
Рис.5 Место реакции в цикле Кребса показано красным цветом.
KEGG Orthology
База ортологов KEGG содержит в себе кластеры ортологичных генов. Ортологи - это гомологичные гены, разошедшиеся в процессе эволюции, т.е. имеющие общего предка. В базе KEGG ген приписывается к определённому кластеру ортологов вручную или с помощью биоинформатических методов. В цикле Кребса выберем реакцию, катализируемую более чем одним набором ортологов, она показана на рисунке 6 розовым. Это реакция превращения малата в оксалоацетат с помощью фермента малатдегидрогеназы (R00342). Она катализируется 3мя рядами ортологичных белков: K00025 (190 белков, из них 16 в Swiss Prot), K00026 (554 белка, из них 18 в Swiss Prot), K00024 (1727 белков, из них 478 в Swiss Prot).
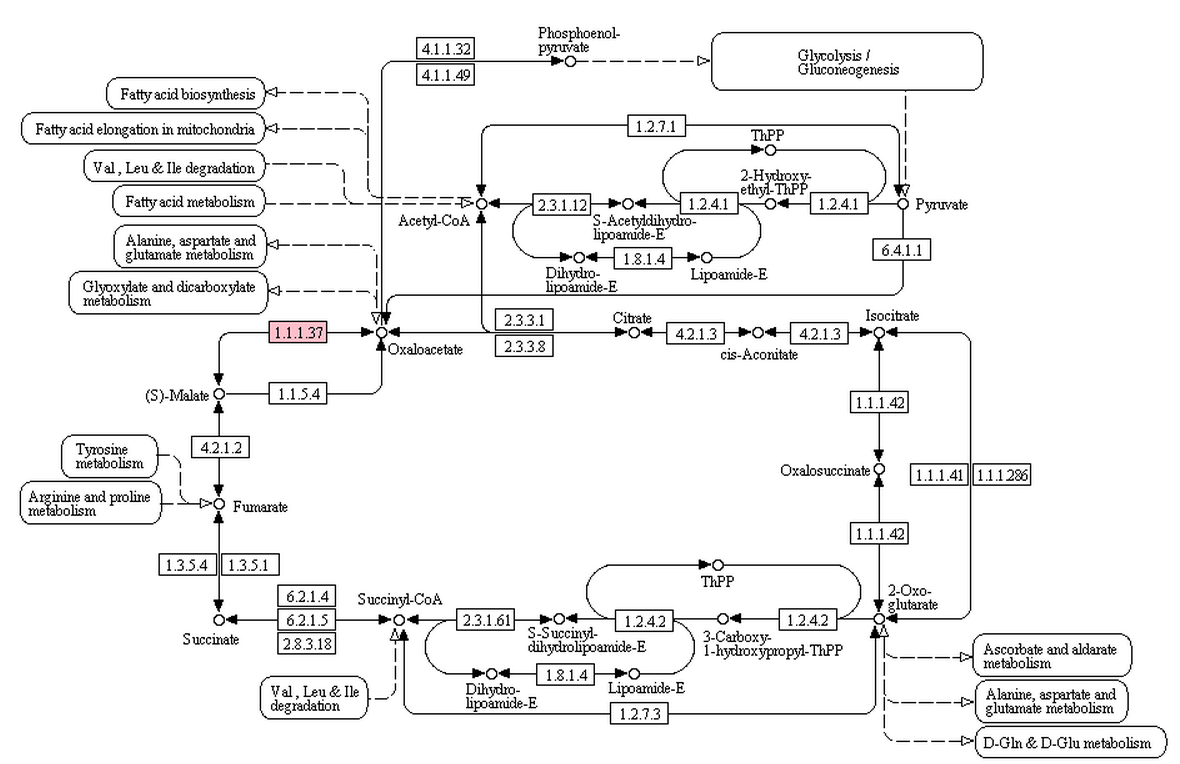
Рис.6. Реакция превращения малата в оксалоацетат, катализируемая тремя рядами ортологичных белков, в цикле Кребса.
Возьмём 16 аннотированных белков из K00025 (их последовательности можно скачать здесь), 18 аннотированных белков из K00026 (последовательности ), 50 аннотированных белков из K00024 (последовательности). Если судить по названиям аннотированных белков, то K00024 - малатдегидрогеназы бактерий, K00025 - эукариот, K00026 - митохондриальные, пероксисомальные и хлоропластные изоформы малатдегидрогеназ. Построим выравнивания этих белков с помощью MUSCLE (фаста-файл, проект Jalview). Фрагмент выравнивания представлен на рисунке 7. Белки выровнялись хорошо: видно, что есть много консервативных позиций. У некоторых белков на N конце виден сигнал митохондриальной или пластидной локализации. В целом С-хвост более вариабельный, ближе к N концу больше консервативных позиций.
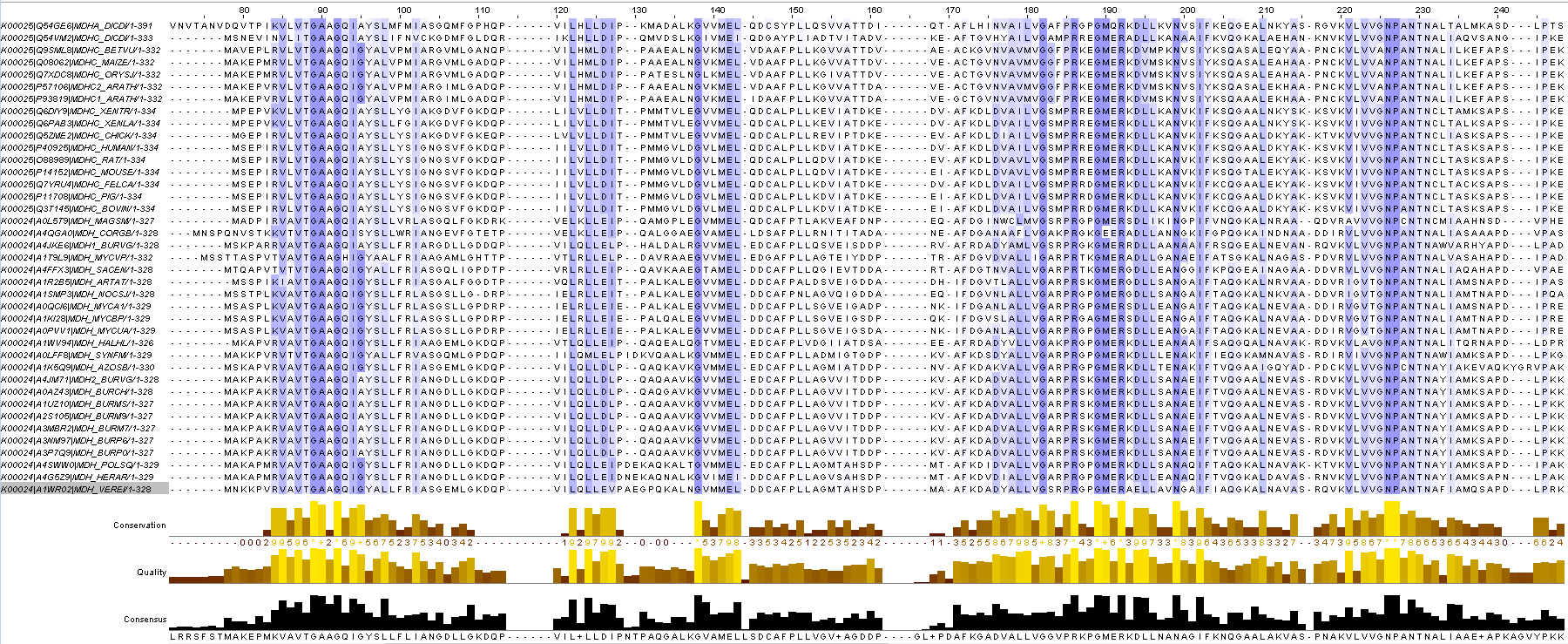
Рис.7 Фрагмент множественного выравнивания белков из трёх разных ортологических рядов малат-дегидрогеназ.
Филогенетическое дерево было построено с помощью MEGA (алгоритм Neighbour-joining). Оно представлено на рисунке 8. Как видно, ортологические ряды K00025 и K00026 действительно выделяются как отдельные клады, однако в ортологическом ряде K00024 есть гены, которые более близки генам из K00025, и гены, которые более близки ортологическому ряду K00026. Если бы эти ортологические ряды были действительно обособлены друг от друга, наблюдалось бы чёткое разделение на 3 клады. Возможно, часть генов из K00024 стоит отнести к K00025, а другую часть - к K00026. Однако, так могло получиться из-за того, что из ортологического ряда K00024 было взято намного больше белков, чем из рядов K00026 и K00025. Далее было построено дерево с 100 бутстреп-репликами, чтобы подтвердить достоверность ветвей. Как видно из рисунка 9, все ветви данного дерева достоверны.
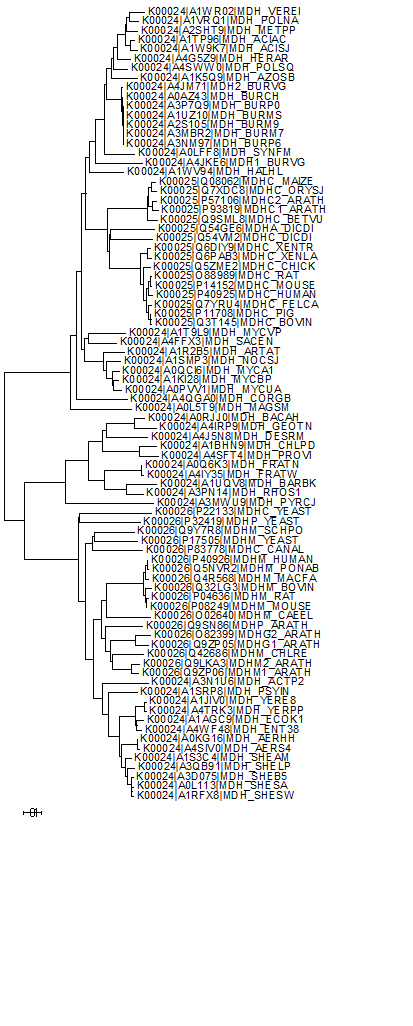
Рис.8. Филогенетическое дерево, построенное в программе MEGA, по выравниванию 16 генов ортологического ряда K00025, 50 генов ортологического ряда K00024 и 18 генов ортологического ряда K00026.
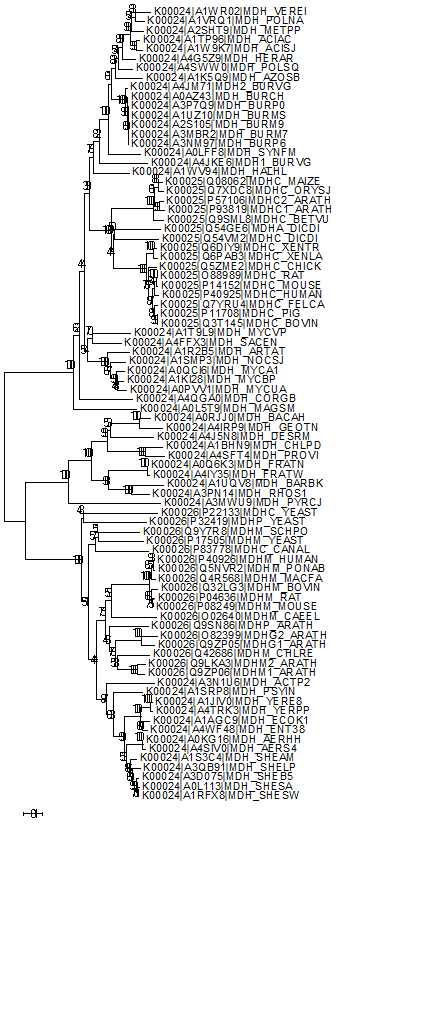
Рис.9. Филогенетическое дерево, построенное в программе MEGA, по выравниванию 16 генов ортологического ряда K00025, 50 генов ортологического ряда K00024 и 18 генов ортологического ряда K00026 c применением бутстрепа.