Редактирование названий последовательностей
Цифрой "3" начинаются последовательности, принадлежащие организмам, которые имеют трехдоменную архитектуру (Glyco_hydro_42M, Glyco_hydro_42, Glyco_hydro_42C); цифрой "2" - двухдоменную (Glyco_hydro_42M, Glyco_hydro_42).
В таблице 1 приведены сокращения таксонов:
| Таксон | Сокращение |
| Actinobacteria | A |
| Firmicutes | F |
| Proteobacteria | P |
| Bacteroidetes | B |
Построение филогенетического дерева
Филогенетическое дерево (рис. 1) построено по методу Neighbor-Joining, который не предполагет молекулярные часы и не выдает длины ветвей.

Рисунок 1. Дерево, построенное по методу Neighbor-Joining.
Можно заметить, что трехдоменные и двухдоменные архитектуры образуют две большие клады. Вероятнее всего, что все три домена изначально имелись у общего предка. Также можно предположить, что произошли как минимум три делеции С-концевого домена. Во-первых, при разделении на две вышеупомянутые клады. Во-вторых, произошла независимая делеция в бактерии 2_P_H3LIH8. И наконец, делеция у общего предка бактерий 2_F_G4H8S7 и 2_F_F3AWV0. Также происходили независимые возникновения доменов в бактериях 3_P_Q2K6V9 и 3_P_A0AXA6. Метод бутстреп подтверждает достоверность дерева (рис. 2), исключая ошибки построения. Следует также отметить, что конвергентная эволция маловероятна из-за большой длины последовательности домена (59 аминокислотных остатков).
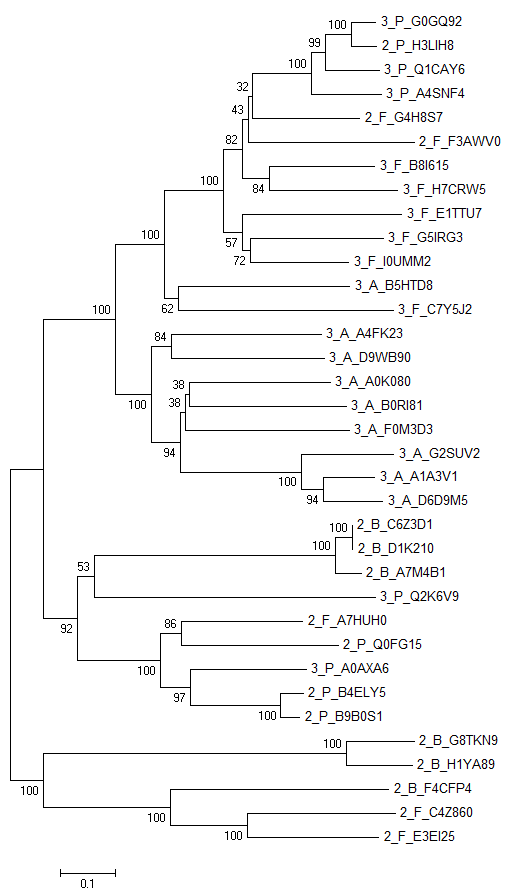
Рисунок 2. Дерево, построенное по методу Neighbor-Joining с использование бутсреп анализа.