В данной работе исследуется особенности генома патогенной бактерии Mycoplasmopsis pulmonis – длины белков, закодированных в ее геноме, нуклеотидный состав кодирующих последовательностей, а также особенности организации генов в геноме.
Систематическое положение изучаемого объекта[1]:
Домен Bacteria
Царство Bacillati
Филум Mycoplasmatota
Отряд Mycoplasmoidales
Семейство Metamycoplasmataceae
Род Mycoplasmopsis
Вид Mycoplasmopsis pulmonis
Mycoplasmopsis pulmonis - бактерия, относящаяся к филуму Mycoplasmatota, отряду Mycoplasmoidales, являюящаяся патогенной и вызывающей
респираторные инфекции у крыс и мышей[2]. Как и все остальные представители Mycoplasmatota, данная бактерия имеет своеобразное строение
клеточных покровов - у нее отсутствует клеточная стенка, покровы представлены исключительно липидной мембранной, вследствие чего
Mycoplasmopsis pulmonisspan является облигатным внутриклеточным паразитом эукариотических организмов. Паразитирует данная микоплазма на
млекопитающих, преимущественно на грызунах, однако есть свидетельства о инфицировании ею людей, что приводило к пневмонии и усилению
других респираторных заболеваний[3].
Так как данная микоплазма инфицирует преимущественно грызунов она является удобной моделью для изучения микоплазменных респираторных
инфекций и микоплазм в целом[2].
В силу внутриклеточного облигатного паразитизма, Mycoplasmopsis pulmonis, как и другие микоплазмы демонстрирует крайне значительную
редукцию генома, как с точки зрения размера – ее геном составляет 963 879 пар оснований, так и с точки зрения числа генов – 782 кодирующие
последовательности, что значительно меньше по сравнению со свободноживущими бактериями. GC - состав генома данной бактерии составляет 26,6% [2].
Исследование было выполнено с помощью методов электронных таблиц (S1)
Исследование было выполнено с помощью методов электронных таблиц (S2)
Исследование было выполнено с помощью методов электронных таблиц (S3)
Для предсказания оперонов использовались электронные таблицы (S4), а также авторский скрипт на языке Python в Google Colab (S5). Как критерий оперона были взяты два и более CDS каждый из которых отстает от предыдущего не более чем на 3 нуклеотида (допускаются пересечения)
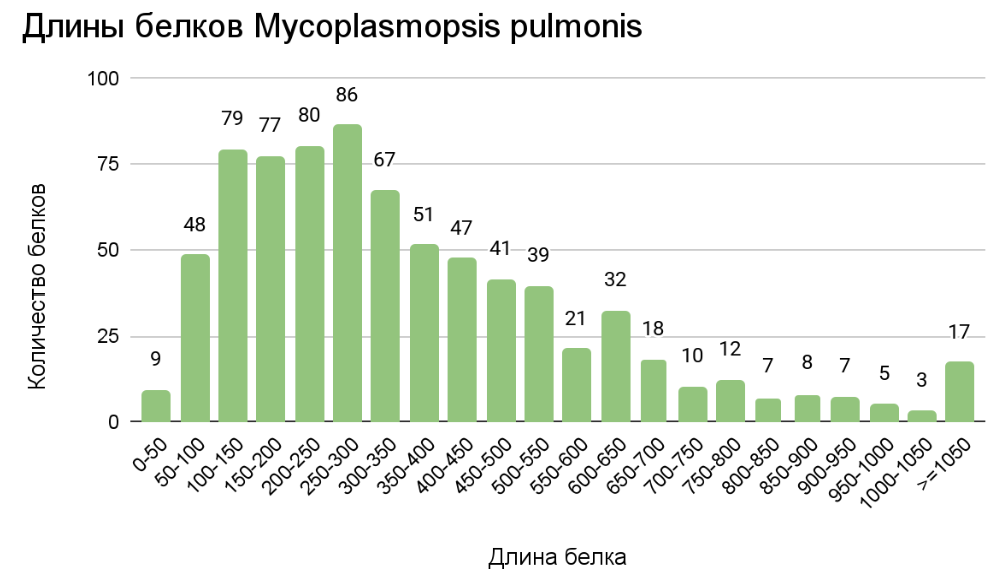
С целью анализа размеров белковых продуктов, закодированных в геноме бактерии M. pulmonis была построена столбчатая диаграмма (рис. 1), отражающая распределение
длин белков
Наибольшее число белков расположилось в кармане 250-300 аминокислот, однако, но также мы можем наблюдать относительное “плато” в распределении на
участке 100 - 300 аминокислотных остатков, белки же длинной более 1000 аминокислотных остатков крайне редки. Медианной длиной является 302 аминокислотных
остатка, самым маленьким белком является 50S рибосомальный белок L36 длиной 37 аминокислотных остатков самым большим – Gli521, структурный белок поверхностного
аппарата скольжения длиной 4279 аминокислотных остатков.
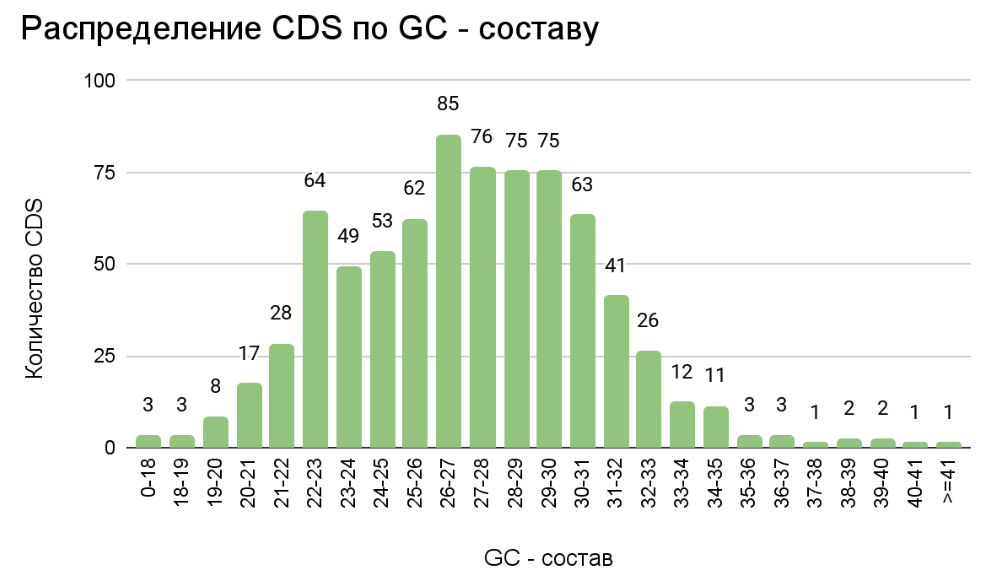
С помощью столбчатой диаграммы (рис. 2) было проанализировано распределение CDS Mycoplasmopsis pulmonis по GC – составу. Анализ гистограммы укрепляет
представление о геноме M. pulmonis, как о геноме типичного внутриклеточного паразита, отражая крайне малое содержание GC в геномных последовательностях.
Процент GC соответствующий наибольшему числу CDS является 26-27% . Однако, мы можем наблюдать дополнительный пик в районе 22-23%. При дополнительном анализе
было выяснено, что повышенное количество CDS в этом диапазоне содержания GC обуславливается содержанием в геноме 17 копий мобильного элемента IS1138B семейства
IS3. Данный мобильный элемент внутри себя кодирует транспозазу и, скорее всего, является активным. Можно предположить, что данный мобильный элемент не потерял
свою активность в результате отбора благодаря паразитическому образу жизни M. Pulmonis и, следовательно, пониженной конкуренции. Медианному значению GC состава
CDS соответствует 27,12%. Самый высокий пик и медианное значение, как и ожидалось, близки к среднему по геному - 26,6%.
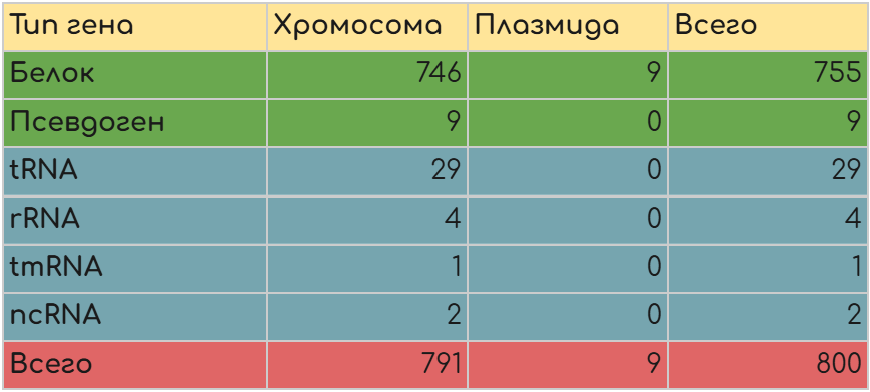
С целью количественного анализа распределения генов различных типов по репликонам генома M. Pulmonis была построена таблица (Таблица 1).
Были подсчитаны количества генов белков, РНК каждого типа, а также количество псевдогенов для всех репликонов M. pulmonis.
На плазмиде данной бактерии находится всего 9 белок-кодирующих генов, все остальные гены (РНК, псевдогены, другие белок-кодирующие) расположены на хромосоме.
Среди 9 генов, располагающихся на плазмиде 7 кодируют белки с неисследованной функцией, оставшиеся два гена tig кодируют триггер-фактор - рибосомальный шаперон,
помогающий сворачиваться новосинтезированным белкам, а также обладающий пролин-изомеразной активностью, что ускоряет свертывание белков [4].
Это позволяет сделать осторожное предположение, что белки на плазмиде (в.т.ч. остальные 7 с неизученной функцией) составляют функциональную группу
трансляционно-ассоциированных “помощников” фолдинга белка, однако, данное предположение требует дополнительного анализа последовательности, структуры и
гомологов данных белков.
С помощью данных о начале и конце генов была составлена таблица предполагаемых оперонов (S6) включившая в себя 38 гипотетических оперонов. В результате функционального анализа найденных оперонов были выделены некоторые, представляющие на взгляд автора наибольший интерес (Таблица 2).
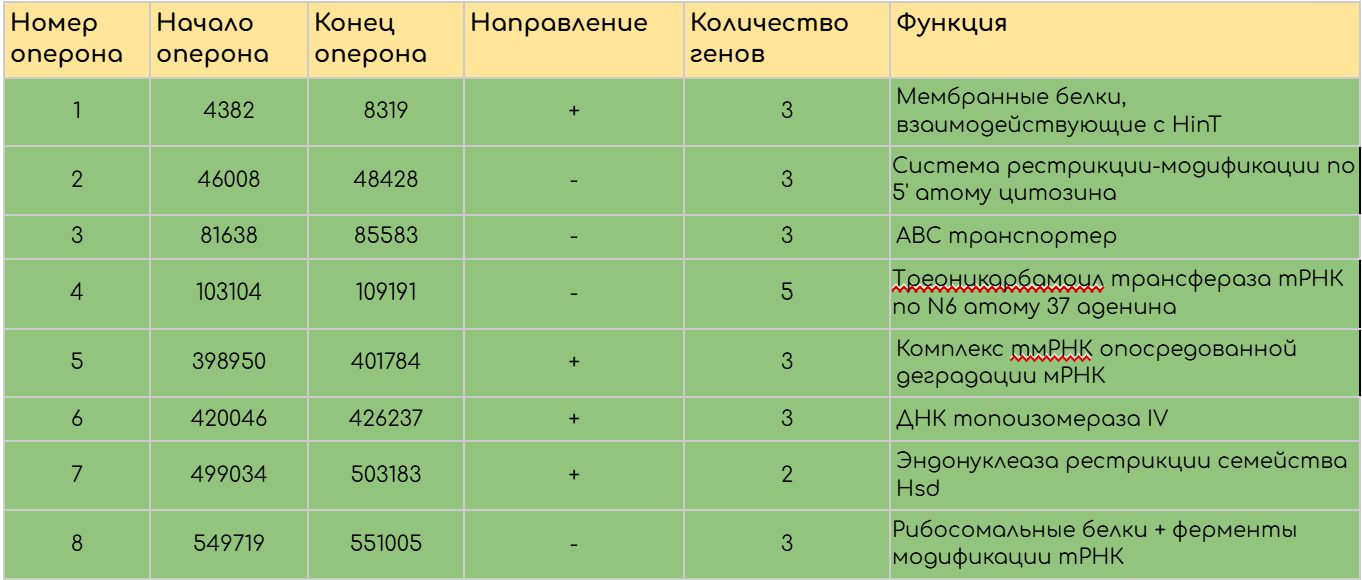
Проведенные исследования, в особенности анализ оперонов, а также более ранние работы других исследователей открывают простор для дальнейшего изучения генома бактерии Mycoplasmopsis pulmonis. Интересным направлением изучения, на взгляд автора, является исследование регуляции системы рестрикции – модификации Hsd. Данная система интересная тем, что ее экспрессия регулируется за счет инверсии генов в локусе. Функциональность и регуляция данной системы была установлена, однако, рекомбиназы, осуществляющей регуляцию найдено не было. Предполагается [2], что регуляция осуществляется за счет рекомбиназы локуса поверхностных антигенов vsa, регулируемого сходным образом. Последовательность, с которой связывается данная рекомбиназа установлена и носит название vrs-бокса[7]. Поиск данной последовательности в опероне Hsd мог бы опровергнуть гипотезу или привести к ней новое доказательство. Также на основании соседей по оперону можно предположить функции гипотетических белков, после чего, с помощью построения моделей структур данных белков и поиска их гомологов у других видов подтвердить или опровергнуть гипотезу о их функциональной принадлежности. Это позволит расширить представления о регуляции экспрессии генов, а также о составе протеома Mycoplasmopsis pulmonis.
Выражаю благодарность
Также все сопроводительные материалы можно найти на моей wiki странице в разделе Сопроводительные материалы