








Работа с KEGG Pathway ↑
Для работы я выбрала путь деградации жирных кислот (fatty acid degradation). Данный путь представлен на сайте KEGG:
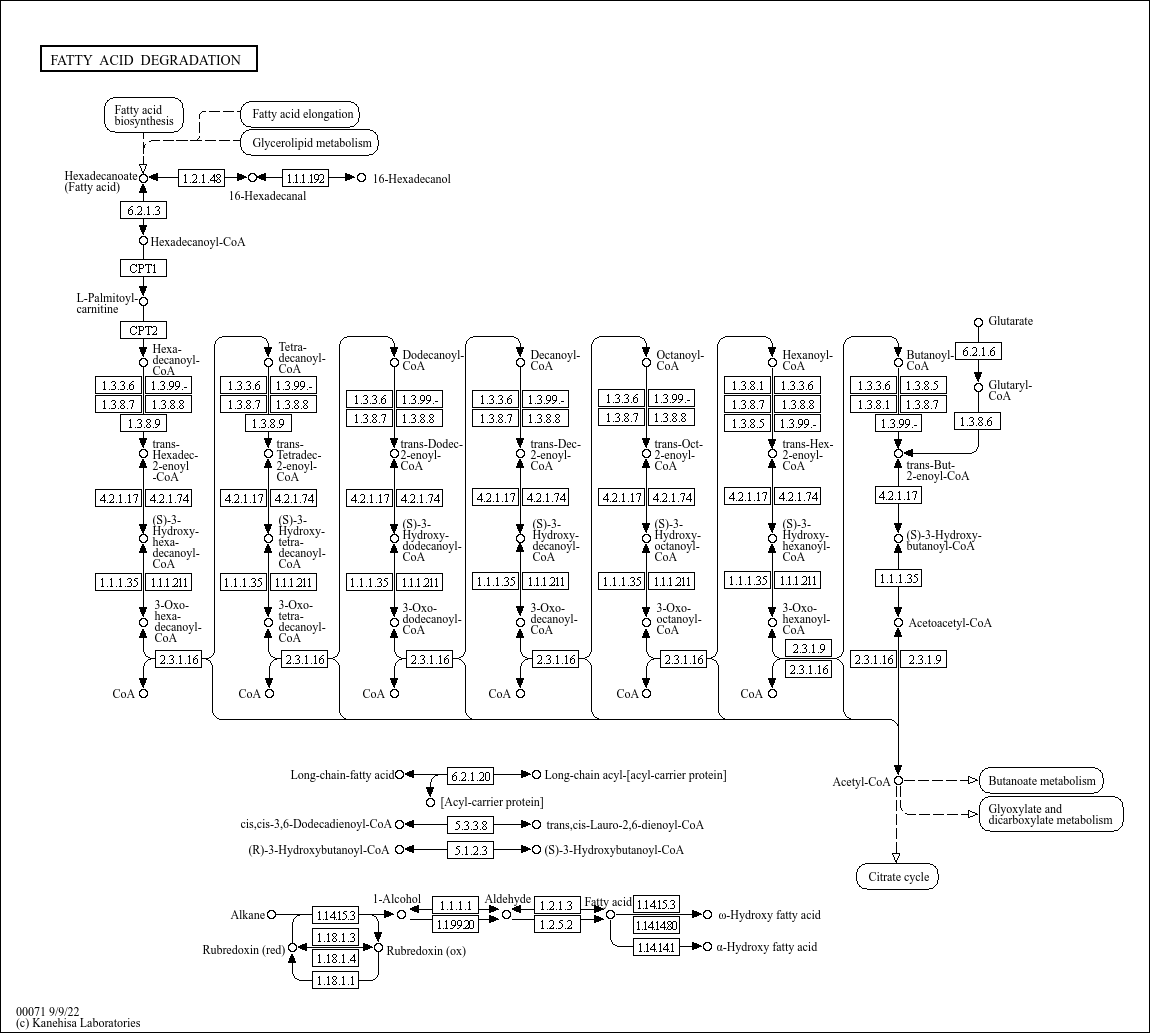
Здесь
хорошо представлен путь деградации жирных кислот. Синтезированные кислоты (на схеме - гексадеканоат, анион пальмитиновой кислоты)
вначале присоединяются к коэнзиму А (ацил-CoA-синтетаза, 6.2.1.3) с образованием RC(O)SCoA. Затем они должны быть перемещены из цитозоля
в митохондриальный матрикс. S-CoA заменяется на молекулу карнитина, в таком состоянии кислота проходит через мембрану в матрикс, где
карнитин сменяется обратно на S-CoA (карнитин О-ацилтрансферазы 1 и 2).
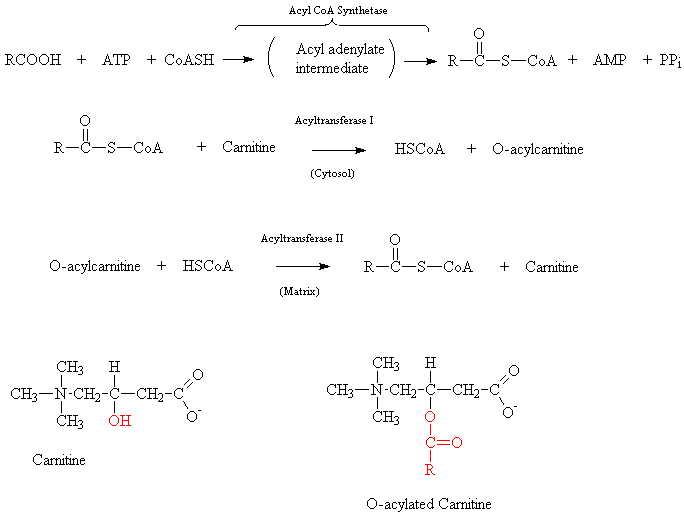
Рис.1.
Дальше идет само β-окисление, проходящее в 4 этапа:- Окисление FAD
- Гидратация
- Окисление NAD+
- Тиолиз CoA
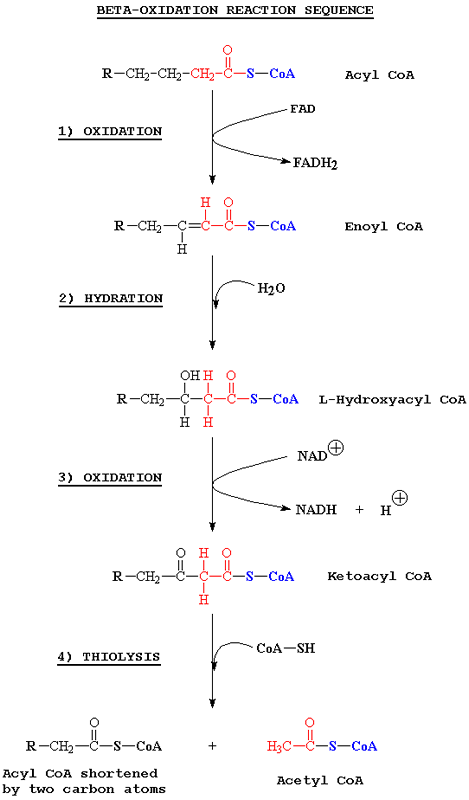
Рис.2. β-окисление жирных кислот
В итоге от кислоты отщепляется 2 углерода со стороны головы. Образовавшийся Ацетил-CoA может далее использоваться в цикле Кребса, а NADH и FADH2 - в окислительном фосфорилировании.
По данным KEGG, путь деградации жирных кислот связан со следующими путями:
- Входящие пути (обеспечивают жирные кислоты):
- Исходящие (используют образовавшийся Ацетил-CoA):
- Побочные входящие:
Чтобы понять, насколько распространен и консервативен этот путь, я выбрала три организма - бактерию Vibrio cholerae O395, архею Sulfolobus islandicus Y.N.15.51 и помидор (Solanum lycopersicum). Ферменты всех стадий основной части цикла (собственно β-окисления) присутствуют у томата и холерного вибриона, хотя стадии окисления FAD у растения и бактерии это делают альтернативные ферменты (1.3.3.6 и 1.3.8.7 у томата и 1.3.99 у вибриона). У обоих есть ацил-CoA синтетаза, но нет карнитин О-ацилтрансфераз. В случае бактерии это понятно, т.к. ей не нужно транспортировать кислоту через митохондриальную мембрану, а как справляется с этим помидор, мне не совсем понятно. У грибов тоже нет карнитин О-ацилтрансфераз, но они есть у всех животных.
У архей, однако, с деградацией жирных кислот все плохо: из всех ферментов присутствуют ацетил-CoA-C-ацетилтрансфераза (2.3.1.9), производящая тиолиз CoA, но она участвует во многих других метаболических путях. У данной археи есть также ацил-CoA синтетаза, но ее нет у многих других архей. Если залезть в путь биосинтеза жирных кислот, то у Sulfolobus там тоже не найдется большей части ферментов. Можно сделать вывод, что археи не занимаются синтезом и деградацией жирных кислот.
Работа с KEGG Reaction ↑
Кроме путей и ферментов, в базе KEGG хранятся биохимические реакции. Например, запись R03857 содержит реакцию Lauroyl-CoA + FAD <=> 2-trans-Dodecenoyl-CoA + FADH2.
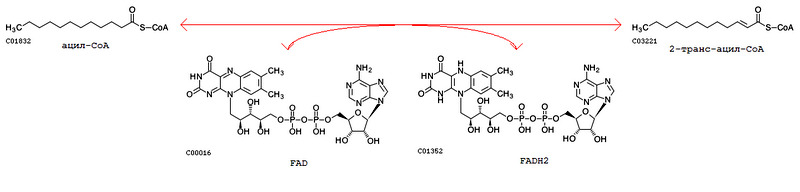
Рис.3.
Подобных реакций в данном метаболическом пути 7, т.к. отдельные реакции записаны для кислот разной длины. Вот их список:
R01279 R03990 R03857 R04754 R03777 R04751 R01175С помощью кнопки User Data Mapping можно покрасить эти реакции в метаболическом пути, например, R01279#FF0000 покрасит рекцию R01279 красным. Вот метаболический путь со всеми реакциями окисления NAD выделенными красным:
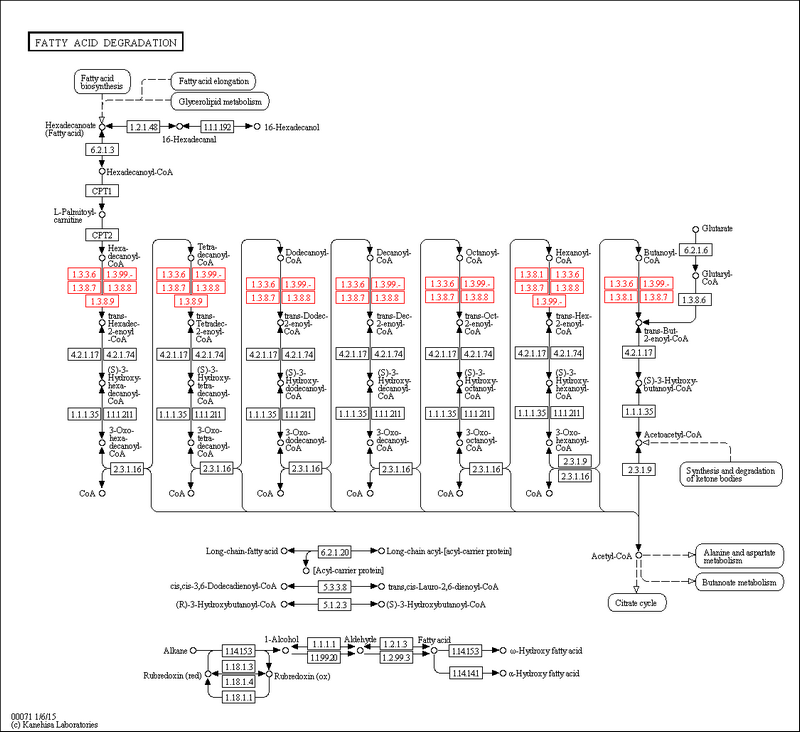
Рис.4. Реакции дегидрирования ацил-CoA
KEGG Orthology ↑
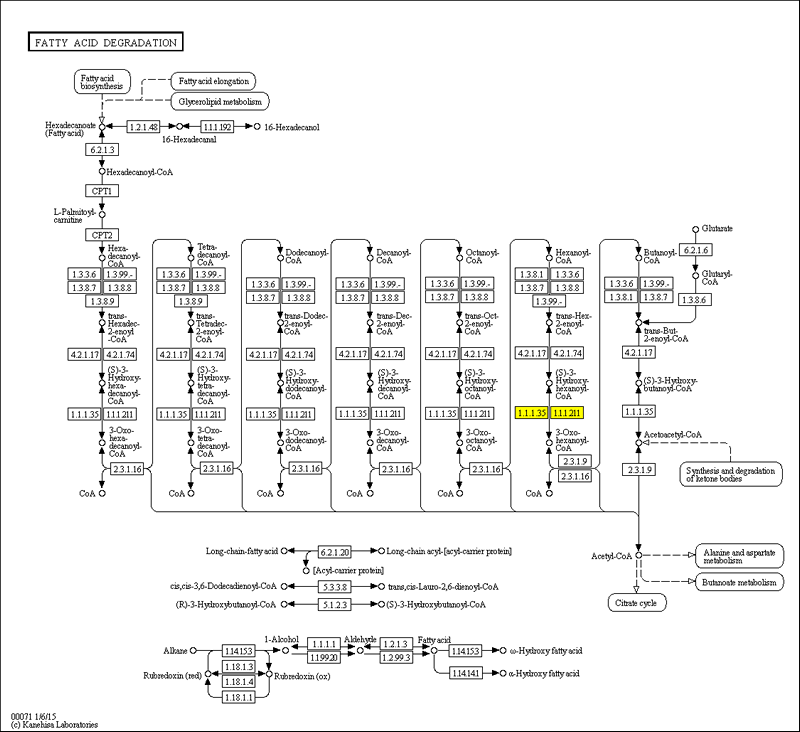
Рис.5. Реакция R04748: 3-гидроксиацил-CoA дегидрирование
Я выбрала реакцию R04748, катализируемую ферментом 3-гидроксиацил-CoA дегидрогеназой (еноил-CoA-гидратазой) (1.1.1.211). Для этой реакции существуют 2 ортологических ряда белков: K07515 и K10527 [x]. В первом из них 68 белков, во втором - 74.
Последовательности белков из этих рядов были скачаны и выровнены сервисом MUSCLE. Проект JalView с выравниванием можно скачать здесь: [x]. Белки из обоих рядов хорошо выровнены. По выравниванию программой MEGA было построено дерево методом Neighbor-joining со 100 бутстрэп-репликами.
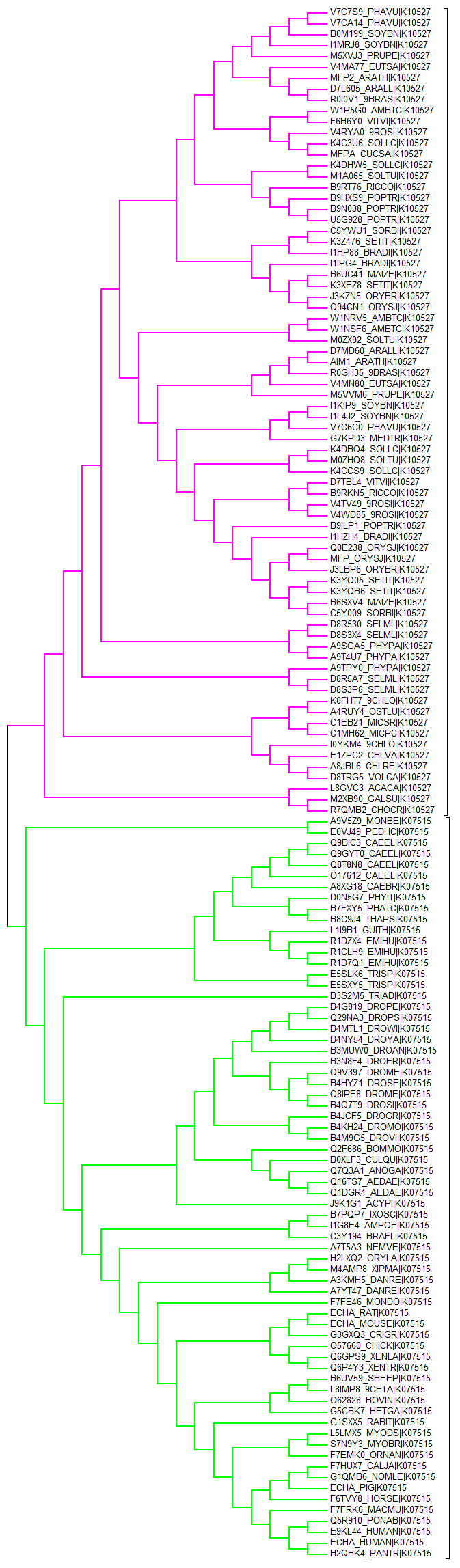
Радует, что дерево четко распадается на 2 клады: первая - гомологический ряд K10527, вторая - K07515. Поддержка бутстрепом ветви с K07515 - 99, т.е. ветвь надежная. Рисунок того же дерева с длинами ветвей и бутстрепом
Я посмотрела на некоторые белки, плохо выровненные с остальными, например A7T5A3_NEMVE|K07515, R1CLH9_EMIHU|K07515, E5SLK6_TRISP|K07515 и L8GVC3_ACACA|K10527 - у них всех есть большие гэпы в нескольких консервативных блоках. На рисунке с длинами ветвей они обозначены синим. L8GVC3_ACACA действительно "сидит" в одиночестве на довольно длинной ветви, R1CLH9_EMIHU сидит на не очень длинной ветви вместе с двумя "соседями" из того же организма, E5SLK6_TRISP тоже расположен вместе с "соседом", A7T5A3_NEMVE не выделяется длиной ветви. Можно сделать вывод, что длинные гэпы на месте некоторых консервативных участков мало влияют на длину ветви, а влияет скорее качество выравнивания там, где эти участки все же есть (т.к. у "соседей" этих гэпов не было, но выравнивание в других местах было схоже). С другой стороны, если посмотреть на одиноко сидящий белок на длинной ветви, например K4C3U6_SOLLC|K10527 (желтая ветвь), можно заметить, что качество выравнивания у него в целом не очень, хотя консервативные участки все на месте. Вывод: при построении деревьев совпадения важнее гэпов.