recombination signal sequence (сигнальная последовательность рекомбинации)
Носитель сигнала
12-RSS и 23-RSS
Кому адресован
ферментативный комплекс RAG1/RAG2
Предназначение — как должен реагировать адресат
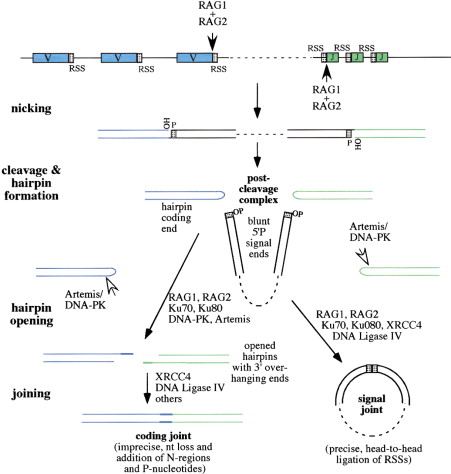
белки RAG узнают сигнал и вносят одноцепочечный разрыв между сегментами V, D и J и их фланкирующими последовательностями RSS
Сила сигнала
Белки RAG должны случайным образом связываться с одной из RSS между V-сегментами и с другой — между J-сегментами. При этом не должно быть такого, что в какой-то области комплекс не связался ни с одной RSS, иначе антитело не получится. Так что сигналы должны быть довольно сильными, а раз все RSS одинаковы по силе, выбор осуществуляется случайным образом
RSS делят на 12-RSS и 23-RSS по длине спейсера. Сплайсинг может происходить только между одним сегментом, фланкированным 12-RSS, и другим сегментом, фланкированным 23-RSS, как гласит правило 12/23. Для оптимальной активности RAG необходимы взаимодействия как с консервативным гептамером, так и с нонамером. 12-RSS и 23-RSS связываются с одним и тем же RAG. Было показано, что белок группы с высокой подвижностью HMGB1 стимулирует ДНК-связывающую активность RAG, предположительно индуцируя изгиб RSS. RSS распознается обеими субъединицами RAG1 в димере RAG, при этом начальная часть в основном распознается инсерционным доменом (ID) и РНКаза H-подобным доменом (RNH) одной субъединицы, а более дистальная часть распознается RNH, ДНК-связывающим доменом и С-концевым доменом (CTD) симметричной субъединицы. Множенственные взаимодействия наблюдаются в первых трёх позициях гептамера, чем объясняется идеальная консервативность последовательности этих нуклеотидов в разных геномах. При этом основание первого нуклеотида выворачивается из дуплекса. Связывание одного RSS индуцирует конформационные изменения в обоих мономерах RAG1, и это может способствовать образованию парных комплексов 12-RSS и 23-RSS. Когда 12-RSS связывается как гептамером, так и нонамером с в HMGB1-изогнутой конформации, димер NBD наклоняется в сторону более короткого 12-RSS, оставляя тот же самый димер NBD со способностью связывания только с 23-RSS, также в HMGB1-изогнутой конформации. И наоборот, когда 23-RSS связан, димер NBD отклоняется в сторону от более длинного 23-RSS, оставляя тот же самый димер NBD со способностью взаимодействовать только с 12-RSS. Также каталитический остаток в активном центре находится на спирали, которая взаимодействует с RSS. Таким образом, связывание RSS может также индуцировать каталитические конформации в RAG1.
После внесения разрыва белками RAG образовавшаяся 3’-ОН группа атакует фосфодиэфирную связь в противоположной цепи. На концах кодирующих сегментов образуются ДНК-шпильки, которые узнаются белками Ku70/Ku80, а Artemis разрезает шпильки (несиквенсспецифично). Далее концы кодирующих сегментов соединяются по тому же механизму, что и при негомологичном соединении концов при репарации ДНК. Так остаются только определённые V, D и J сегменты. Перед этим терминальная дезоксинуклеотидил трансфераза (TdT) достраивает несколько нуклеотидов (до 10-20) случайным образом, что дает дополнительную вариабельность. Вырезанный участок, содержащий сигнальные последовательности RSS, замыкается с образованием кольцевидной структуры, известной как рекомбинационное вырезанное кольцо.
Рис. 1. Схема V(D)J рекомбинации и расположения кодирующих сегментов и RSS.
Литература:
1. Mark S. Schlissel "Does Artemis End the Hunt for the Hairpin-Opening Activity in V(D)J Recombination?" doi:10.1016/S0092-8674(02)00694-3
2. Heng Ru, Melissa G. Chambers, Tian-Min Fu, Alexander B. Tong, Maofu Liao, and Hao Wu "Molecular Mechanism of V(D)J Recombination from Synaptic RAG1-RAG2 Complex Structures" doi: 10.1016/j.cell.2015.10.055
3. Uthayashanker R. Ezekiel,Tianhe Sun,Grazyna Bozek &Ursula Storb "The Composition of Coding Joints Formed in V(D)J Recombination Is Strongly Affected by the Nucleotide Sequence of the Coding Ends and Their Relationship to the Recombination Signal Sequences" https://doi.org/10.1128/MCB.17.7.4191
4. Craig H. Bassing, Frederick W. Alt, Maureen M. Hughes, Margaux D'Auteuil, Tara D. Wehrly, Barbara B. Woodman, Frank Gärtner, J. Michael White, Laurie Davidson and Barry P. Sleckman "Recombination signal sequences restrict chromosomal V(D)J recombination beyond the 12/23 rule" Nature 405, 583–586 (2000) https://doi.org/10.1038/35014635
5. Dale A. Ramsden, Tanya T. Paull & Martin Gellert "Cell-free V(D)J recombination" Nature 388, 488–491 (1997) https://doi.org/10.1038/41351
6. Alastair L. Smith, James N. F. Scott, Joan Boyes "The ESC: The Dangerous By-Product of V(D)J Recombination" https://doi.org/10.3389/fimmu.2019.01572
Сервис для поиска сигнала
Для поиска положения сайтов рекомбинации можно использовать разные программные методы, реализованные в пакете программ RDP v. 3.34: RDP (Recombination detection program); Geneconv (Statistical Tests for Detecting Gene Conversion); BootScan (Bootstrap-based detection of recombination breakpoints); Chimaera (Maximun mismatch chi-square); 3Seq (Method for inferring mosaic structure in sequence triplets); Siscan (Sister-scanning), MaxChi (Maximum chi-square). Программы выполняют статистическую оценку результатов. Результат считают достоверным, когда значение р было равным или меньше 0,05.
Программа обнаружения рекомбинаций (RDP) — это компьютерная программа, используемая для анализа данных о нуклеотидных последовательностях и выявления признаков генетической рекомбинации. Помимо применения большого количества различных методов обнаружения рекомбинаций, он также реализует различные методы построения филогенетических деревьев и recombination hotspot тесты.
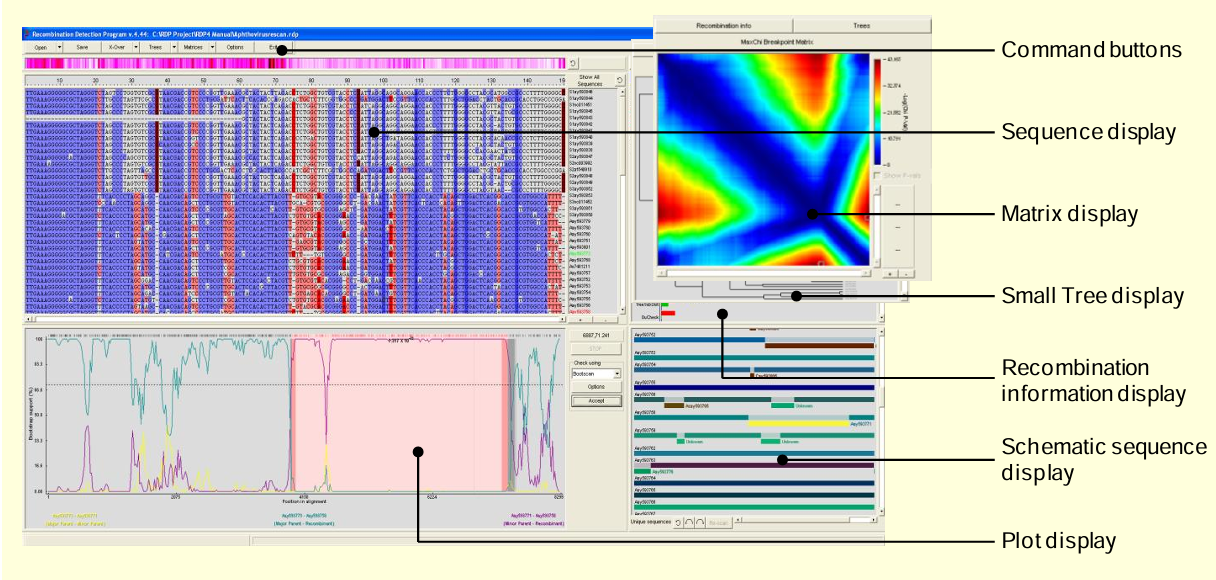
Рис. 2. Основные компоненты интерфейса RDP.
После загрузки файлов последовательностей в любом из различных форматов с помощью кнопки "Открыть" на панели командных кнопок, нажатие кнопки "X-Over" начнет автоматическое исследовательское сканирование на рекомбинацию с текущими установленными параметрами анализа (их можно просмотреть, нажав кнопку "Опции"). Доступ к различным визуализациям филогенетических деревьев и матриц набора данных можно получить через стрелки рядом с кнопками "Деревья" и "Матрицы". Можно переключаться между отображениями дерева, матрицы и информации о рекомбинации с помощью кнопок над ними.
В разделе «Анализ последовательностей с помощью» можно выбрать алгоритмы обнаружения, которые будут выполняться по протоколу RDP.
Рис. 3. Настройка RDP.
Выбераем «Выполнить» → «Выполнить полное исследовательское сканирование рекомбинации». После завершения анализа можно исследовать каждое предлагаемое событие рекомбинации различными способами. В правом нижнем углу можно увидеть отображение последовательности схем. Цветные блоки в этом окне графически представляют идентифицированные события рекомбинации. Каждая последовательность представлена в виде блока, а под ним цветные полосы представляют потенциальные события рекомбинации, происходящие в этой последовательности.
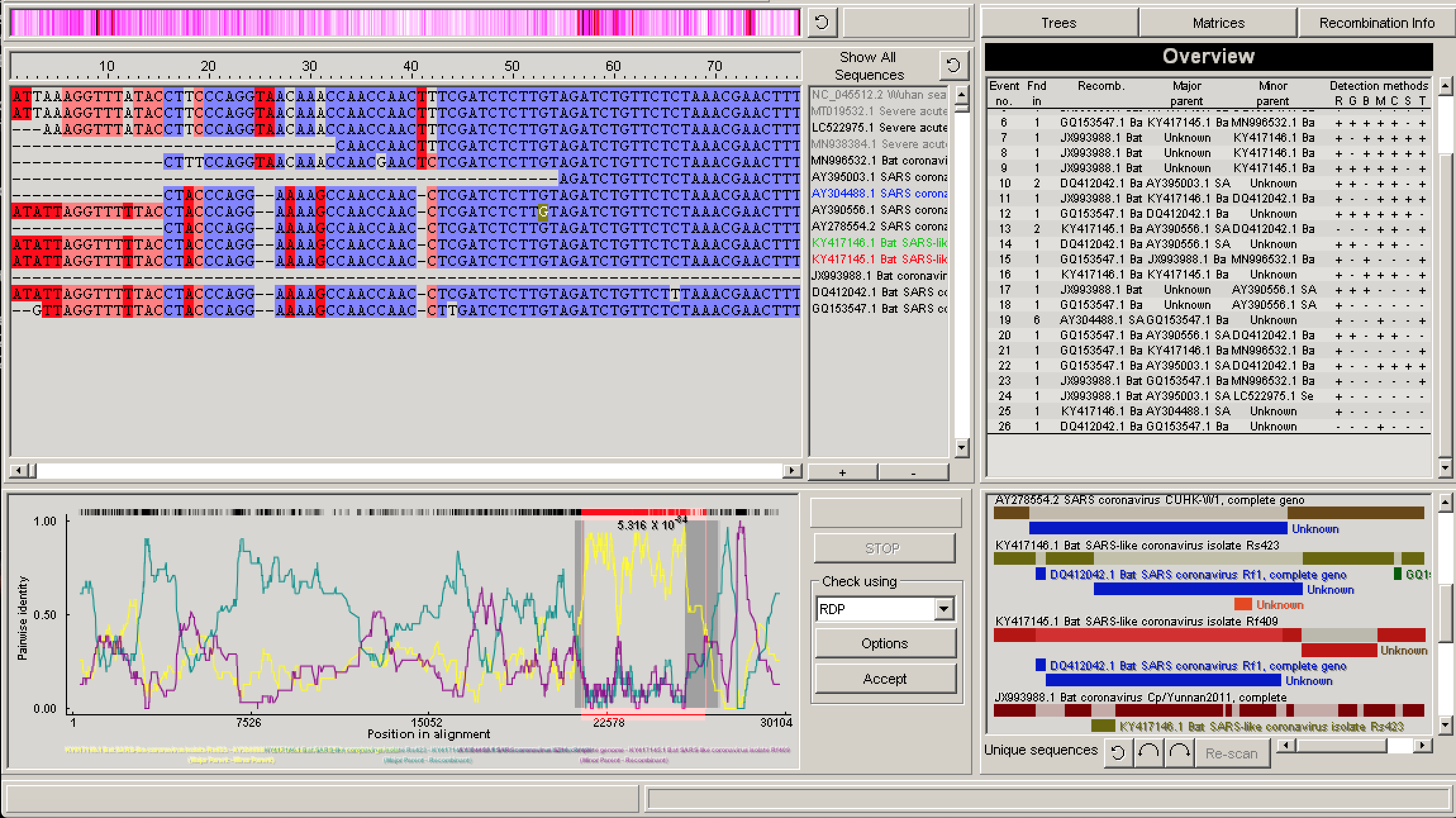
В правом верхнем углу находится информационное окно «Рекомбинация», в котором перечислены все идентифицированные события рекомбинации, наиболее вероятные родительские последовательности и методы обнаружения, которые положительно определили событие рекомбинации.
Рис. 4. Обзор анализа RDP.
При выборе события рекомбинации, щелкнув одно из событий в окне «Информация о рекомбинации» или один из цветных блоков на экране последовательности схемы, можно затем щелкнуть «Информация о рекомбинации», чтобы получить более подробную информацию о событии.
Источники
1. Recombination Detection Program (RDP) user manual http://web.cbio.uct.ac.za/~darren/RDP4Manual.pdf
2. Darren P Martin, Ed Rybicki RDP: Detection of recombination amongst aligned sequences doi:10.1093/bioinformatics/16.6.562