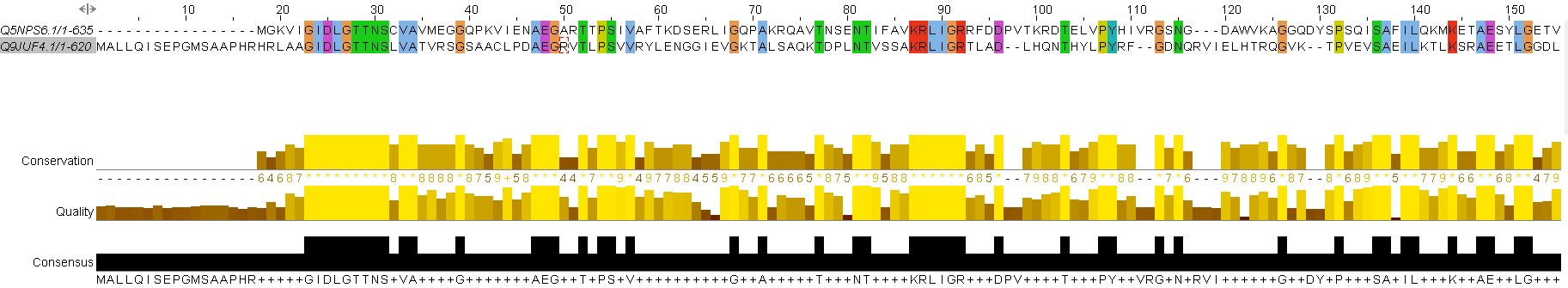
Характеристики выравниваний:
Матрица весов - EBLOSUM62. Выравнивания, полученные при штрафах по умолчанию, вышли идентичными. Я решила поправить ситуацию с помощью задания более жёстких штрафов – 10.0 за открытие и закрытие инделя, 10.0 – за его удлинение, 10.0 – за концевые гэпы, 0.5 – за концевые индели.
1. Снимки первых позиций выравниваний:
a) глобального
b) локального

2. Снимки последних позиций выравниваний:
a) глобального

b) локального
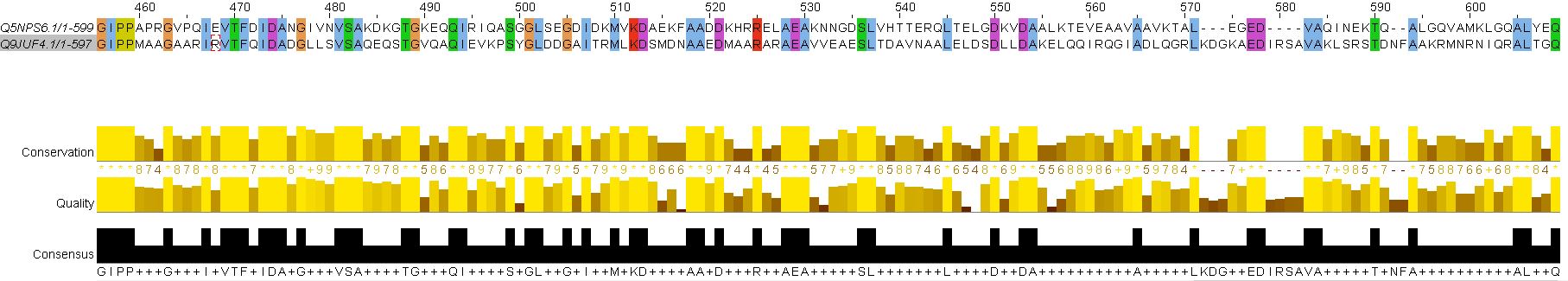
Одно из отличий глобального выравнивания от локального – программа локального выравнивания обрезала сильно различающиеся участки в начале и в конце последовательностей и начала выравнивание с полностью консервативных позиций. В случае глобального выравнивания эти участки заполнялись гэпами. В связи с этим длина глобального выравнивания заметно больше, чем длина локального.
Сравнение параметров глобального и локального выравнивания:
| Длина выравнивания | Консервативные позиции | Функционально консервативные позиции | Колонки с гэпами | Число инделей | ||||
| число | % | число | % | число | % | |||
| Глобальное выравнивание | 666 | 263 | 39.5 | 373 | 56.0 | 77 | 11.6 | 15 |
| Локальное выравнивание | 622 | 418 | 62.6 | 506 | 76.3 | 31 | 5.0 | 12 |
| ID белков | Длина выравнивания | Консервативные позиции | Функционально консервативные позиции | Колонки с гэпами | Число инделей | |||
| число | % | число | % | число | % | |||
| RIBF_CORAM, O15922_LEIMA |
47 | 13 | 27.7 | 19 | 40.4 | 1 | 2.1 | 1 |
| RIBF_CORAM, NHAA_RHOER |
44 | 14 | 31.8 | 18 | 40.9 | 0 | 0 | 0 |
| RIBF_CORAM, Q89GW5_BRADU |
19 | 3 | 15.8 | 8 | 42.1 | 0 | 0 | 0 |
| RIBF_CORAM, TIP21_ARATH |
40 | 12 | 30 | 19 | 47.5 | 1 | 2.5 | 1 |
| RIBF_CORAM, EXOS7_HUMAN |
16 | 8 | 50 | 10 | 62.5 | 0 | 0 | 0 |
Поскольку в данных выравниваниях я использовала те же строгие штрафы, что и в случае гомологичных белков, программа локального выравнивания сильно обрезала белки во избежание лишних гэпов. Поскольку негомологичные белки сильно отличаются, гэпов в выравниваниях должно было образовываться огромное колическтво.
Вот, к примеру, снимок первого из пяти полученных выравниваний.
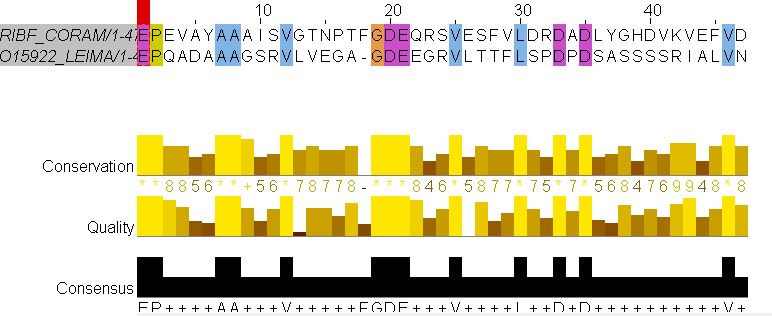
Для сравнения приведу фрагмент того же выравнивания, но со штрафами по умолчанию - 10.0 за открытие и закрытие инделя, 0.5 за его удлинение.

Видно, что данное выравнивание заметно длиннее и содержит большее количество гэпов.
Стоит отметить, что локальное и глобальное выравнивание очень похожи между собой. Единственное найденное мною отличие - глобальное выравнивание на один а.к.о. длиннее локального.
Множественное выравнивание отличается от двух других сильнее. Оно совпадает с ними вплоть до 285 позиции. Во множественном выравнивании в белке DNAK_ARTS2 этой позиции соответстует гэп, стоящий напротив глутаминовой кислоты (Е) в белке DNAK_SALAI. В глобальном и локальном выравниваниях они расположены наоборот. Далее различия замечены только ближе к концу. С 667 позиции в строках DNAK_SALAI парных выравниваний начинается большой индель из 5 гэпов, а участок AQA выровнен с участком SAA (672-674 позиции). В случае множественного выравнивания этот индель отстутствует, участок AQA вместе с GAA сразу сопоставлен участку PAGAES последовательности DNAK_ARTS2 (667-672) - в двух других выравниваниях GAA смещены длинным инделем вправо и стоят напротив GSE (704-706). Также на позиции 733 в случае парных выравниваний глутаминовая кислота (Е) в DNAK_ARTS2 соответстует лизину (К), в то время как в множественном лизин смещен гэпом на соседней 732 позиции.
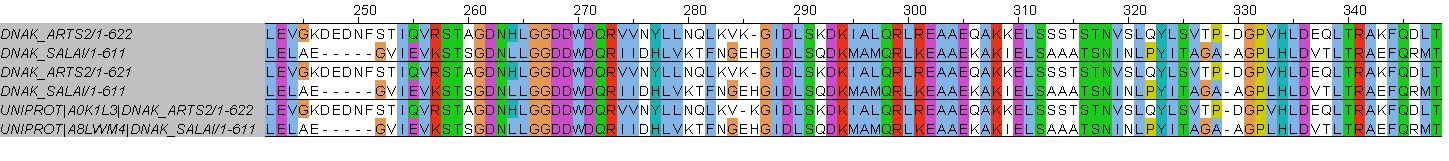
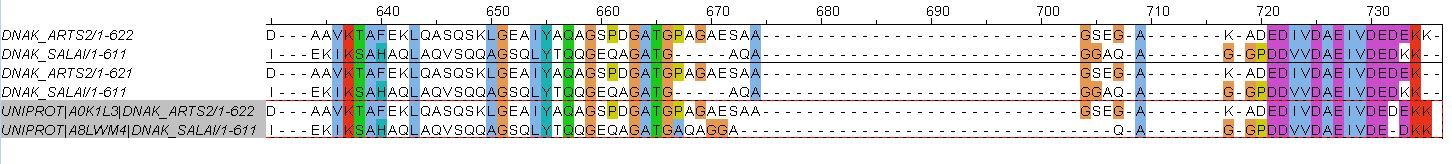
Я считаю наиболее правдоподобным выравнивание, полученное из множественного выравнивания 10 практикума. Оно скорее будет верно отражать гомологичные аминокислотные остатки, так как при построении опирается на большее количество последовательностей.