Практикум 2. Алгоритмы филогенетической реконструкции
Получение выравнивания цитохромов B
Я решил построить выравниевание цитохромов B на kodomo для выбранных
ранее десяти представителей Teleostei. Был создан файл cyb.list.
С помощью команды seqret @cyb.list cyb.fasta был получен файл с последовательностями
цитохромов B для выбранных мной организмов из банка Swiss-prot. Далее было проведено выравнивание алгоритмом muscle:
muscle -align cyb.fasta -output cyb-alignment.fasta.
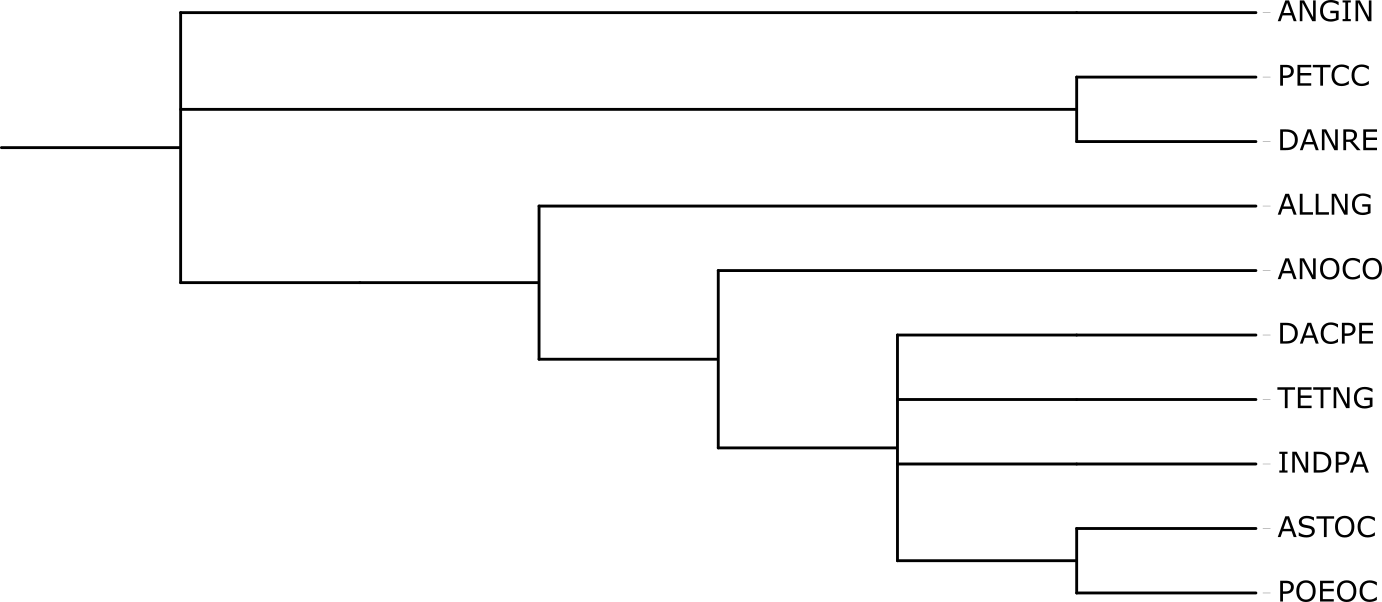
Рис.1. Филогенетическое дерево для некоторых видов инфракласса Teleostei, соответствующее реальной таксономии.
Для укоренения всех трёх деревьев я выбрал ветвь, ведущую к ANGIN, поскольку согласно реальной таксономии ANGIN образует отдельную монофилетическую ветвь, и я использовал её в качестве внешней группы. Это позволило получить бифуркационное дерево, в котором корень расположен между ANGIN и остальными таксонами.
Реконструкция дерева программой FastME
С помощью скрипта Python выравнивание было переведено из формата fasta в phylip-relaxed и записано в файл cyb.phy.
Деревья по моделям p-distance и MtREV были построены соответственно с помощью команд
fastme -i cyb.phy -o pdistance.tre -pp и fastme -i cyb.phy -o mtrev.tre -pM
FastME MtRev
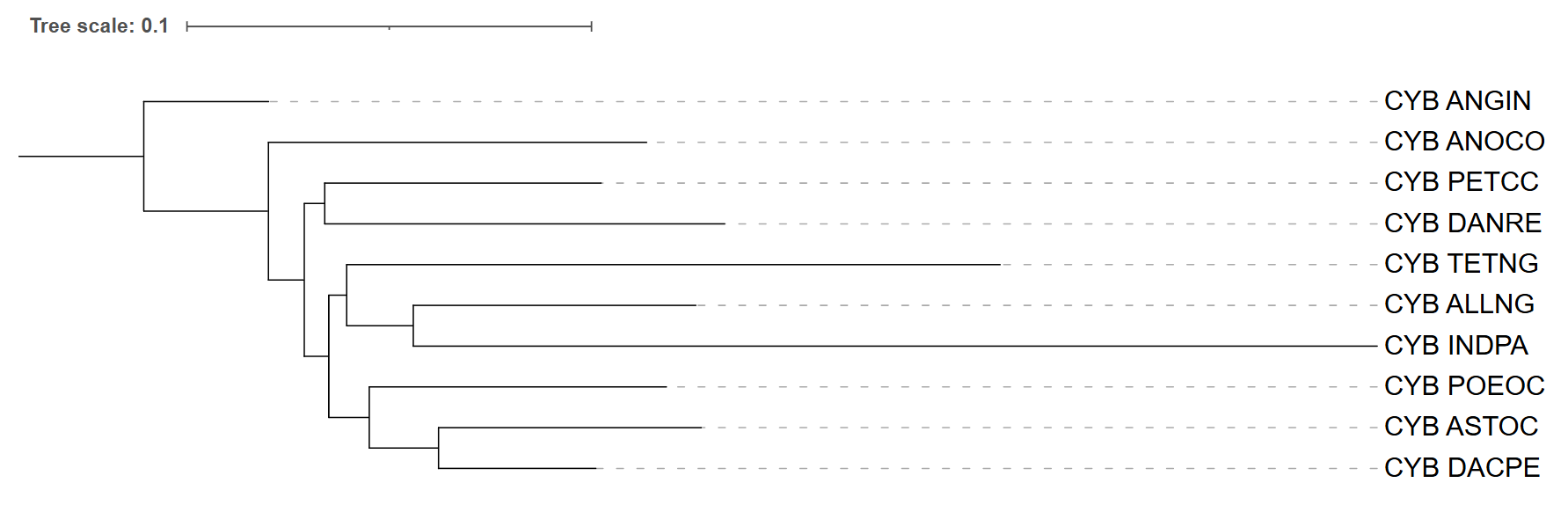
Рис. 2. Дерево FastMe MtRev. Укоренение проведено в ветвь, ведущую к ANGIN.
Сводка:
- Последовательности: выравнивание цитохрома B для 10 видов инфракласса Teleostei.
- Модель эволюции: MtREV (специализированная для митохондриальных белков).
- Алгоритм реконструкции: минимальная эволюция (Minimum Evolution).
- Программа: FastME (запуск с ключом -pM).
- Укоренение: выполнено в ветвь, ведущую к виду ANGIN (использован как внешняя группа на основании того, что в реальной таксономии ANGIN образует отдельную монофилетическую ветвь).
Дерево укоренено на ветви, ведущей к виду ANGIN, который вынесен в качестве внешней группы (это обосновано тем, что в реальной таксономии ANGIN образует отдельную монофилетическую ветвь). Топология имеет следующий вид:
(ANGIN,(ANOCO,((PETCC,DANRE),((POEOC,(ASTOC,DACPE)),(TETNG,(ALLNG,INDPA))))))
В этом дереве ANOCO оказывается базальным ко всем таксонам, кроме ANGIN, что не соответствует таксономии, где ANOCO входит в большую кладу вместе с DACPE, INDPA, TETNG, POEOC и ASTOC, а ALLNG является сестринским к этой группе. Главное совпадение с таксономией – правильное восстановление клады (DANRE, PETCC); эти два вида объединены в монофилетическую группу, как и ожидалось. Также частично верно сгруппированы POEOC, ASTOC и DACPE – они образуют одну подветвь, однако по таксономии DACPE должна быть самостоятельной ветвью, а не объединяться с (POEOC, ASTOC).
Основные расхождения с реальной таксономией:
- ALLNG сгруппирован с INDPA и TETNG (вместо того чтобы быть базальным к кладе (ANOCO, DACPE, INDPA, TETNG, POEOC, ASTOC))
- INDPA и TETNG образуют общую кладу с ALLNG, тогда как по таксономии они являются отдельными ветвями внутри большой клады
- ANOCO занимает неправильное базальное положение.
FastMe p-distance
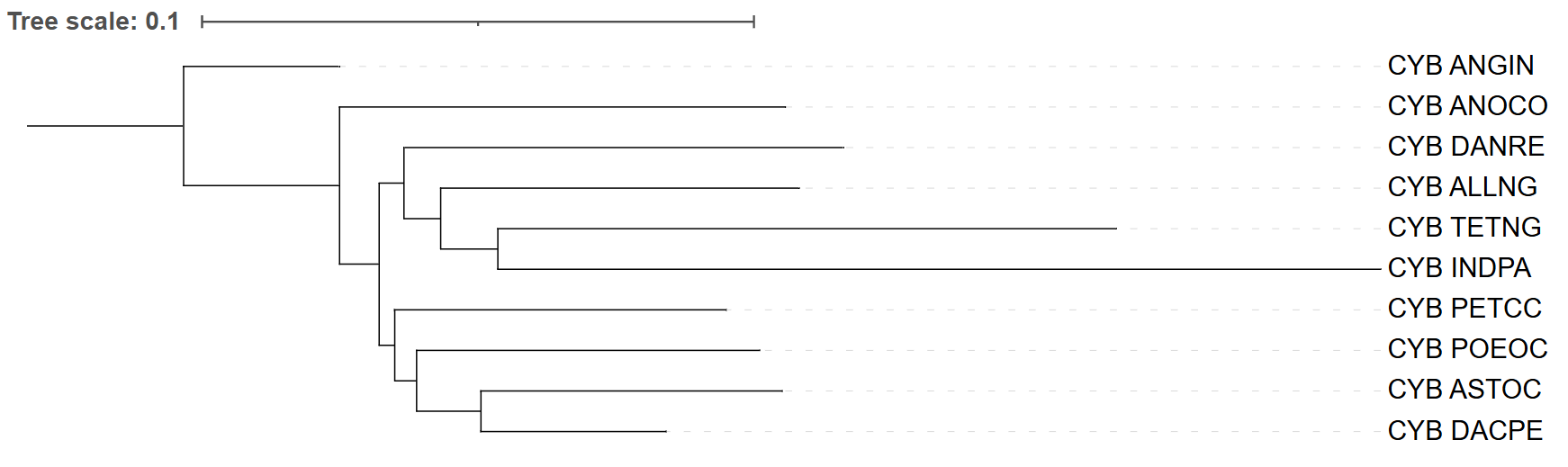
Рис. 3. Дерево FastMe p-distance. Укоренение проведено в ветвь, ведущую к ANGIN.
Сводка:
- Последовательности: то же выравнивание цитохрома B для 10 видов Teleostei.
- Модель эволюции: p‑distance (простые попарные расстояния без коррекции на множественные замены).
- Алгоритм реконструкции: минимальная эволюция.
- Программа: FastME (запуск с ключом -pp).
- Укоренение: аналогично, на ветвь к ANGIN (внешняя группа).
Укоренение выполнено аналогично – на ветви к ANGIN, который также выступает внешней группой. Топология:
(ANGIN,(ANOCO,((PETCC,(POEOC,(ASTOC,DACPE))),(DANRE,(ALLNG,(TETNG,INDPA))))))
В этом дереве ANOCO снова базальна (ошибка, общая с MtREV). Однако здесь гораздо больше расхождений с таксономией: ключевая клада (DANRE, PETCC) полностью разрушена: PETCC оказывается в одной ветви с POEOC, ASTOC и DACPE, а DANRE попадает в группу с ALLNG, TETNG и INDPA. Также все четыре таксона (PETCC, POEOC, ASTOC, DACPE) объединяются в одну монофилетическую группу, тогда как по таксономии PETCC должен быть с DANRE, а DACPE – отдельной ветвью, не сгруппированной с (POEOC, ASTOC).
Вторая большая клада – (DANRE, ALLNG, TETNG, INDPA) – также противоречит таксономии: DANRE должен быть с PETCC, ALLNG – базален ко всей основной группе, а INDPA и TETNG – отдельные ветви. Единственное частичное сходство – наличие подклады (ASTOC, DACPE) внутри ветви с POEOC и PETCC, но это не компенсирует множественные ошибки.
Реконструкция дерева программой IQ-Tree
С помощью команды iqtree -s cyb.phy (с параметрами по умолчанию) дерево было реконструировано. Здесь оно выглядит ближе к систематике, чем деревья, реконструированные программой FastME. ANGIN расположен отдельано, а ASTOC, POEOC и DACPE находятся в отдельной ветви. Остальая топология также варьируется.
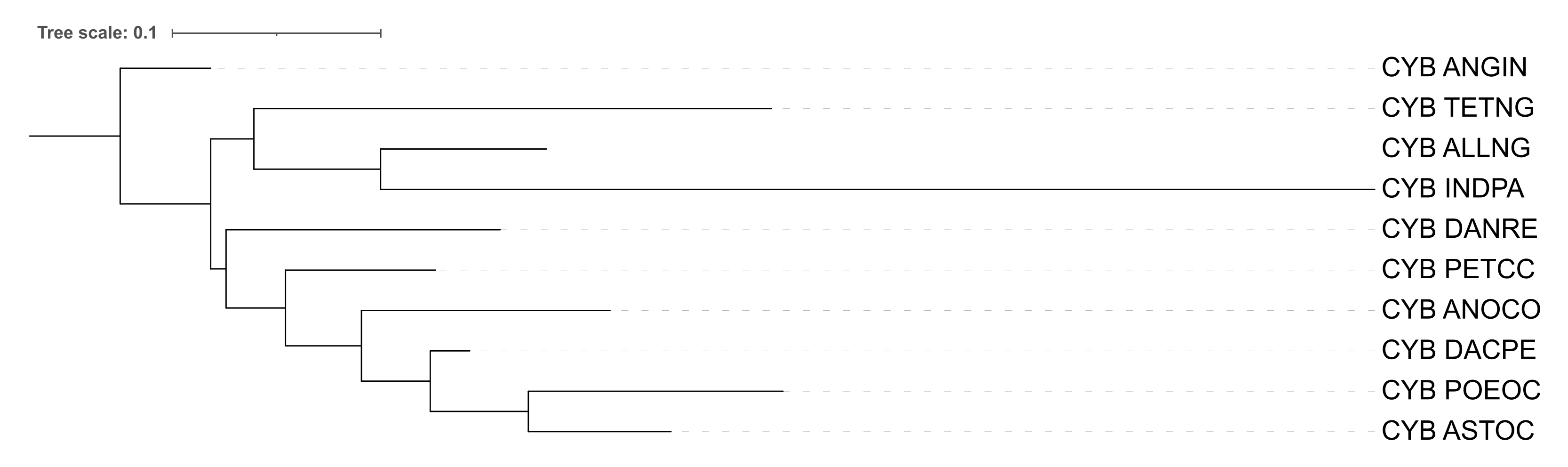
Рис. 4. Дерево IQ-Tree. Укоренение проведено в ветвь, ведущую к ANGIN.
Сводка:
- Последовательности: то же выравнивание цитохрома B для 10 видов Teleostei.
- Модель эволюции: автоматически выбрана программой по критерию BIC – mtZOA+I+G4 (смесь эмпирических митохондриальных матриц с учётом инвариантных сайтов и гетерогенности скоростей).
- Алгоритм реконструкции: максимальная вероятность (Maximum Likelihood).
- Программа: IQ‑TREE 2 (запуск с параметрами по умолчанию, iqtree -s cyb.phy).
- Укоренение: выполнено в ту же ветвь – к ANGIN (внешняя группа).
Дерево укоренено на ветви к ANGIN, выступающему в роли внешней группы. Его топология:
(ANGIN,((TETNG,(ALLNG,INDPA)),(DANRE,(PETCC,(ANOCO,(DACPE,(POEOC,ASTOC)))))))
Сходства с реальной таксономией:
- ANOCO помещён внутрь большой клады вместе с DACPE, POEOC и ASTOC – это соответствует таксономии, где ANOCO входит в основную группу (в отличие от MtREV и p‑distance, где ANOCO был базальным).
- Клада (POEOC, ASTOC) восстановлена правильно (они образуют монофилетическую подгруппу).
- DACPE находится в той же большой кладе, что и POEOC/ASTOC, что частично верно (хотя по таксономии DACPE должна быть самостоятельной ветвью, а не сгруппированной с ними).
Различия с реальной таксономией:
- Клада (DANRE, PETCC) не восстановлена – PETCC оказывается в одной ветви с ANOCO, DACPE, POEOC и ASTOC, а DANRE – отдельно, что противоречит таксономии.
- ALLNG сгруппирован с INDPA, а не является базальным к основной кладе. Кроме того, TETNG оказывается сестринским к (ALLNG, INDPA), хотя по таксономии TETNG и INDPA – отдельные ветви внутри большой клады.
- INDPA и TETNG не занимают правильных независимых позиций.
IQ‑TREE даёт смешанный результат: исправляет ошибку с положением ANOCO, но теряет правильную группировку (DANRE, PETCC), которая была верно восстановлена в MtREV.
Выводы
Ни один из методов не дал полностью правильного дерева, что ожидаемо при использовании единственного белка (цитохром b) и ограниченного набора таксонов. Однако модель MtREV показала наилучший результат, поскольку правильно восстановила монофилетическую группу (DANRE, PETCC) – наиболее важную таксономическую кладу. Модель p‑distance, будучи самой простой, привела к наибольшему числу искажений. IQ‑TREE с автоматическим подбором сложной модели (mtZOA+I+G4) продемонстрировал компромиссный вариант: он улучшил положение ANOCO, но за счёт потери другой критической клады. Это указывает на то, что выбор эволюционной модели существенно влияет на топологию, и даже продвинутые методы не гарантируют полной точности без дополнительных данных (например, большего числа генов или более репрезентативной выборки).