Практикум 7. Трансмембранные белки
1. Информация о выбранном белке
Я работал с данным мне из таблицы белком 6ln7. Он представляет из себя Mg-зависимую АТФ-азу P-типа, использующую энергию АТФ для активного транспорта ионов Ca2+ из цитоплазмы в люмен саркоплазматического ретикулума. Играет ключевую роль в мышечном расслаблении и кальциевом гомеостазе клетки.
Таблица 1. Сводка о белке
| Название (рус.) | Кальциевая АТФаза саркоплазматического/эндоплазматического ретикулума 2 (SERCA2, изоформа) |
|---|---|
| Название (англ.) | Sarcoplasmic/endoplasmic reticulum calcium ATPase 2 (SERCA2b) |
| PDB ID | 6LN7 |
| UniProt AC | P16615 |
| Ген | ATP2A2 |
| Организм | Homo Sapiens |
| Локализация в клетке | Мембрана саркоплазматического ретикулума и эндоплазматического ретикулума (SR/ER-мембрана) |
Последовательность белка (каноническая изоформа 1, 1042 a.a.) была взята из UniProt (P16615).
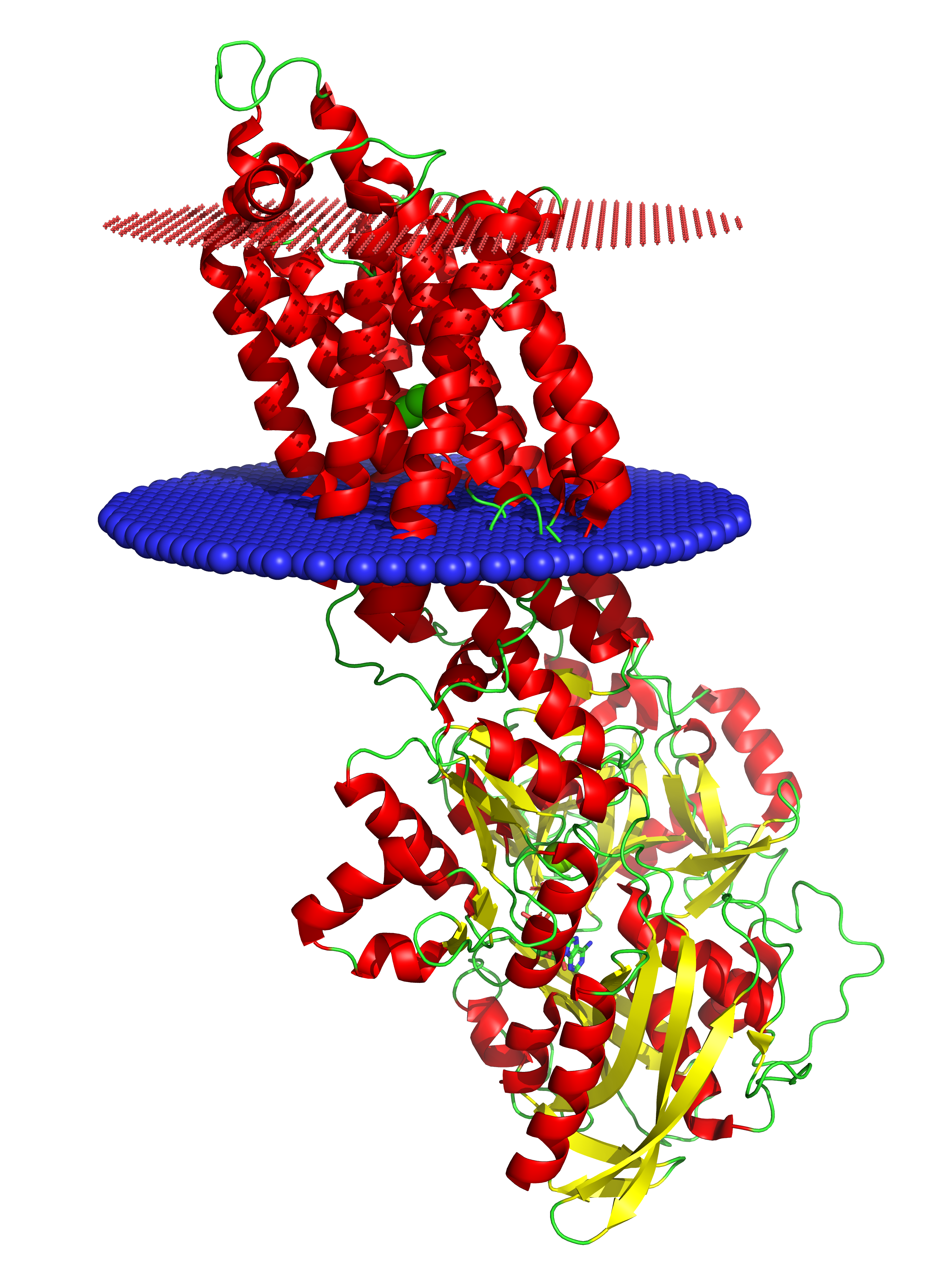
Рис. 1. Трехмерная структура 6LN7. Синим обозначен внутренний липидный слой, красным – внешний.
2. Трансмембранные участки по данным OPM (Orientations of Proteins in Membranes)
Для белка SERCA2 (PDB 6LN7) в базе OPM определены следующие трансмембранные сегменты. Информация получена со страницы записи о 6LN7.
В общей сложности 11 трансмембранных α-спиралей, что типично для P-тип АТФ-аз. Границы определены по положению белка относительно рассчитанной гидрофобной толщины липидного бислоя
Таблица 2. Список трансмембранных участков для 6LN7 согласно базе данных OPM. Цепь A, нумерация согласно канонической последовательности UniProt P16615
| Номер спирали | Начало | Конец | Длина (a.a.) |
|---|---|---|---|
| TM1 | 59 | 79 | 21 |
| TM2 | 85 | 105 | 21 |
| TM3 | 259 | 279 | 21 |
| TM4 | 288 | 313 | 26 |
| TM5 | 758 | 780 | 23 |
| TM6 | 788 | 806 | 19 |
| TM7 | 832 | 853 | 22 |
| TM8 | 895 | 914 | 20 |
| TM9 | 929 | 948 | 20 |
| TM10 | 965 | 986 | 22 |
| TM11 | 1013 | 1029 | 17 |
3. Предсказания трансмембранных участков с помощью DeepTMHMM
Для предсказания использовалась та же каноническая последовательность UniProt P16615 (изоформа 1, 1042 а.о.) = в формате FASTA. Сервис DeepTMHMM был запущен с параметрами по умолчанию (модель Alpha-globular). Результаты работы были сохранены в файле TMRs.gff3
Были определены 11 трансмембранных α-спиралей, столько же, сколько и при использовании OPM.
Таблица 2. Список трансмембранных участков для 6LN7 согласно базе данных OPM. Цепь A, нумерация согласно канонической последовательности UniProt P16615
| Номер спирали | Начало | Конец | Длина (a.a.) |
|---|---|---|---|
| TM1 | 60 | 76 | 17 |
| TM2 | 88 | 108 | 21 |
| TM3 | 256 | 276 | 21 |
| TM4 | 296 | 314 | 19 |
| TM5 | 762 | 782 | 21 |
| TM6 | 787 | 807 | 21 |
| TM7 | 836 | 856 | 21 |
| TM8 | 895 | 915 | 21 |
| TM9 | 930 | 949 | 20 |
| TM10 | 966 | 986 | 21 |
| TM11 | 1018 | 1030 | 13 |
Графическое представление результатов DeepTMHMM
На Рис. 2. показаны апостериорные вероятности принадлежности каждого положения к трём состояниям: Inside (внутри, цитоплазма), Membrane (мембрана) и Outside (снаружи, люмен SR/ER). Отчётливо видны 11 пиков, соответствующих трансмембранным спиралям.
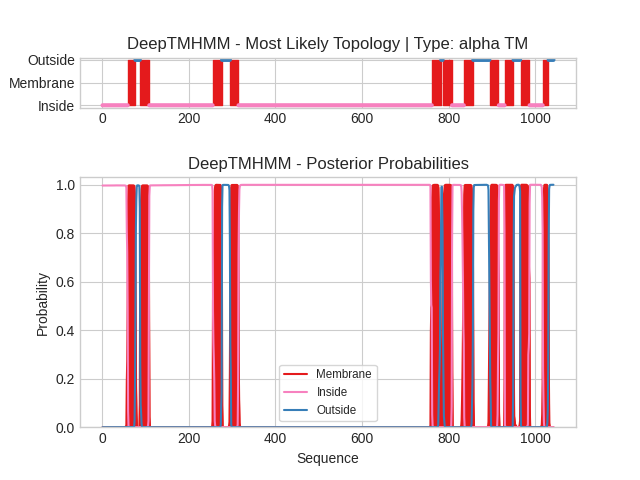
Рис. 2. Предсказание трансмембранных участков белка SERCA2 (P16615) с помощью DeepTMHMM.
4. Качественное сравнение результатов OPM и DeepTMHMM
Оба метода – OPM, основанный на трёхмерной структуре и расчётном позиционировании в липидном бислое, и DeepTMHMM, работающий только с аминокислотной последовательностью, – предсказали одинаковое число трансмембранных спиралей (11). Практически все спирали перекрываются на 80-100% своей длины. Расхождения составляют от 1 до 8 аминокислотных остатков на N- или C-концевых участках спирали. Например:
- TM1: OPM 59–79, DeepTMHMM 60–76 – ядро перекрывается (60–76), OPM считает на 3 остатка длиннее с каждого конца.
- TM4: OPM 288–313, DeepTMHMM 296–314 – перекрывающаяся часть 296–313, концы различаются не более чем на 8 остатков.
- TM9: OPM 929–948, DeepTMHMM 930–949 – идентично по длине, сдвиг на 1 остаток.
Расхождения можно объяснить несколькими факторами:
- Физическая основа методов разная: OPM определяет трансмембранный участок как часть цепи, погруженную в гидрофобное ядро бислоя с учетом точной атомной структуры. DeepTMHMM опирается только на статистические закономерности в последовательностях, выявленных на основе множества белков с известной топологие, и предсказывает положение спирали исключительно на основании последовательности белка.
- Концевые участки спиралей часто содержат амфифильные или полярные остатки, которые находятся на липидно-водой границе. OPM и DeepTMHMM могут по-разному интерпретировать положения этих остатков, считая или не считая их находящимися внутри мембран
- Вследствие погрешности обучения DeepTMHMM может незначительно смещать границы трансмембранных сегментов, если в тренировочныых даных распределение длин определенных спиралей имело систематический тренд.
Несмотря на расхождения, можно сделать вывод, что предсказание глубокого обучения успешно воспроизводит структурные данные о трансмембранной топологии SERCA2, подтверждая надёжность обоих подходов. Все 11 трансмембранных элементов надёжно идентифицируются, а небольшие различия в границах не превышают погрешностей, типичных для методов предсказания мембранной топологии.