Мини-обзор генома бактерии Mycetocola zhujimingii
Mycetocola это род, впервые описанный в 2001 году в составе семейства Microbacteriaceae. В 2019 году была описана бактерия Mycetocola zhujimingii, она была выделена из фекалий Тибетской антилопы (Pantholops hodgsonii)[1]. Данная антилопа обитает на самом высоком нагорье в мире (Цинхай-Тибетском нагорье) и считается почти самым малоизученным парнокопытным животным. Mycetocola zhujimingii это грамположительная, не образующая спор, аэробная бактерия, размерами примерно 0,3– 0,6×0,9–2,2 мкм. Полную таксономию можно видеть в таблице 1.
|
Систематическая группа |
Наименование |
|
Тип |
Actinomycetota |
|
Класс |
Actinomycetia |
|
Порядок |
Micrococcales |
|
Семейство |
Microbacteriaceae |
|
Род |
Mycetocola |
|
Вид |
Mycetocola zhujimingii |
Результаты данного мини-обзора были получены с помощью ряда методов. Для анализа таблицы особенностей бактерии Mycetocola zhujimingii были использованы Google sheets, благодаря данному инструменту были построены различные гистограммы: распределение длин белков (формула =СЧЁТЕСЛИМН), исследование пересекающихся генов, кодирующих белки (формула =ЕСЛИ), исследование расстояний между генами и таблицы распределение генов белков и разных типов РНК по репликонам (формула =СЧЁТЕСЛИМН), общее число различных кодирующих последовательностей и их типов. Исходная таблица особенностей M. zhujimingii была скачана с NCBI и уже с ней впоследствии производились преобразования[2].
Из рисунка 1 можно сделать вывод, что для данной бактерии наиболее характерны белки длиной 200-300 аминокислотных остатков. Также много белков чуть меньшей длины (100-200 аминокислотных остатков) и чуть большей длины (300-400 аминокислотных остатков). Самым длинным(1950 аминокислотных остатков) оказался белок Ig-like domain-containing protein. Белки с иммуноглобулиноподобными доменами часто используются бактериями для клеточной адгезии[3], также подобные белки были найдены у паразитических энтеробактерий[4]. Из данной информации и знания о том, откуда была выделена эта бактерия, можно сделать предположение, что она обитает в кишечнике антилопы (а не была занесена на фекалии другими способами). Самым маленьким белком (32 аминокислотных остатка) оказался AURKAIP1/COX24 domain-containing protein с не до конца ясной функцией.
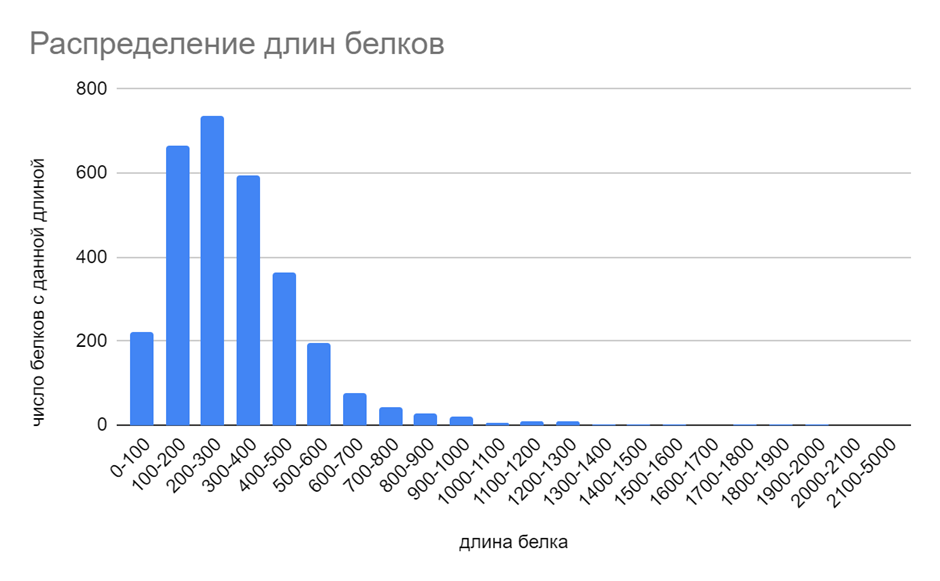
Рис. 1. Гистограмма длин белков.
Таблица 2. Количество генов, кодирующих РНК и белки.
| Gene | 3062 |
| CDS | 3003 |
| tRNA | 47 |
| ncRNA | 2 |
| tmRNA | 1 |
| rRNA | 9 |
Таблица 3. Распределение белков и различных типов РНК по репликонам.
| genomic_accession | seq_type | chromosome | CDS | tRNA | rRNA | ncRNA | tmRNA |
| NZ_CP026949.1 | chromosome | 2996 | 47 | 9 | 2 | 1 | |
| NZ_CP026950.1 | plasmid | unnamed | 37 | 0 | 0 | 0 | 0 |
Из таблицы 3 видно, что все виды РНК закодированы в хромосомном геноме, в то время как в геноме плазмиды закодировано всего 37 белков. Большая часть из них гипотетические, много белков, участвующих в репликации плазмиды и её последующей передаче другим бактериям. Но в глаза сразу бросается ген, кодирующий cobalamin biosynthesis protein CobQ (белок биосинтеза кобаламина). Скорее всего, в биосинтезе кобаламина и заключается основная функция плазмиды. (у некоторых видов из кишечной микрофлоры человека тоже имеется такая способность, что ещё больше нас наталкивает на мысль, что перед нами именно энтеробактерия).
Из рисунка 2 следует, что для данной бактерии наиболее характерны расстояния между генами, кодирующими белки, равные от 800 до 900 нуклеотидов. При этом достаточно часто встречаются расстояния больше (от 900 до 1000 нуклеотидов) и меньше (от 400 до 800 нуклеотидов). В этих промежутках могут располагаться как важные регуляторные последовательности, необходимые для регуляции экспрессии генов, так и гены различных РНК.
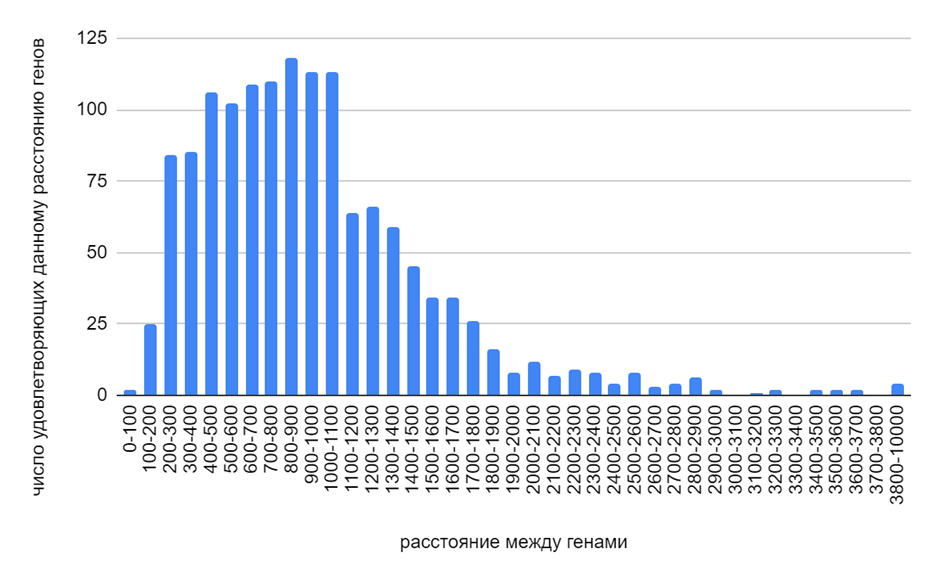
Рис. 2. Гистограмма длин белков M. zhujimingii.
Для Mycetocola zhujimingii наиболее характерны перекрывания генов длиной от 1 до 5 нуклеотидов. При этом наибольшей интерес представдяют гены, котрые перекрываются на куда большие интервалы нуклеотидов. К примеру, ген рекомбиназы/интегразы тирозинового типа перекрывается на 106 нуклеотидов с геном N-терминальной фаговой интегразы, содержащей SAM-домен (в её активном центре так же имеются остатки тирозина). Можно сделать предположение, что когда-то фаг заразил предковую бактерию и встроил свой геном в геном бактерии, последующие мутации могли привести к возникновению из гена фага нового гена со сходной функцией продукта трансляции[5]. Ещё я обратил внимание на 3 последовательно пересекающихся гена (то есть первый ген пересекается со вторым, а второй с третьим). Причём первый ген первый пересекается со вторым на 31 нуклеотид, а второй с третьим на 6 нуклеотидов. 1 ген — УДФ-N-ацетилмурамоил-L-аланин-D-глутаматлигаза. 2 ген — липид II-флиппаза FtsW. 3 ген — УДФ-N-ацетилглюкозамин-N-ацетилмурамил-(пентапептид) пирофосфорил-ундекапренол-N-ацетилглюкозаминтрансфераза. Все 3 гена задействованы в построении клеточной стенки у бактерий [6,7,8]. Поэтому смею предположить, что первые 2 гена могут экспрессироваться как один цистрон, в результате чего FtsW (трансмембранный переносчик) может служить якорем для фермента УДФ-N-ацетилмурамоил-L-аланин-D-глутаматлигаза. В связи продукта 3 гена с FtsW я не конца уверен, ведь пересечение относительно небольшое, но всё же не исключено, что N-ацетилглюкозаминилтрансфераза тоже может закрепляться на мембране благодаря переносчику FtsW.
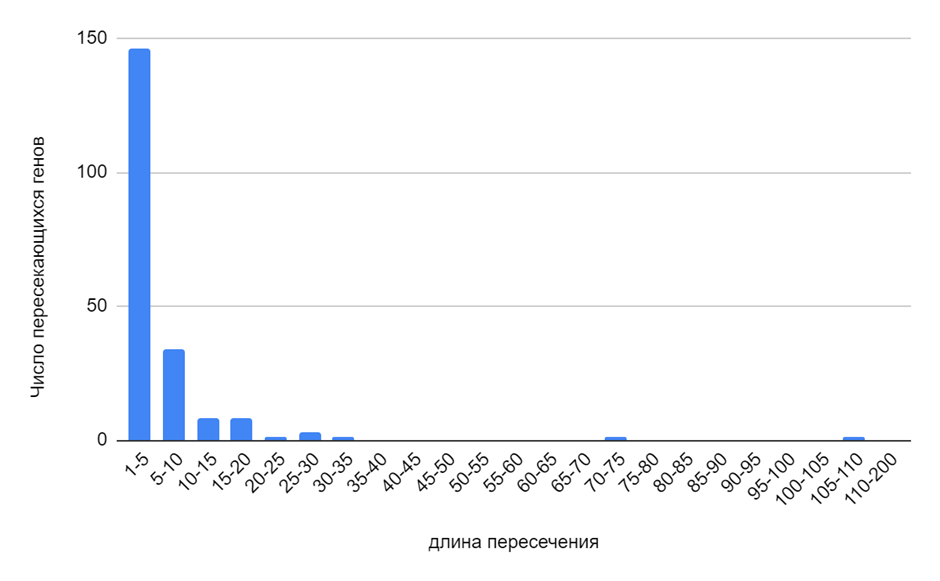
Рис. 3. Распределение интервалов пересечения генов у Mycetocola zhujimingii.