Практикум 3
Задание 1
В данном задании была предсказана вторичная структура тРНК различными способами: путём поиска инвертированных повторов и по алгоритму Зукера с помощью ViennaRNA. Полученные результаты я сравнила с результатами, полученными ранее с помощью программы find_pair пакета 3DNA.
Программа einverted была запущена со следующими параметрами:
Gap: 12
Threshold: 10
Match: 3
Mismatch: -3
В результате работы программы einverted был найден только участок, соответствующий акцепторному стеблю. При уменьшении параметра threshold результат выдачи не менялся.
Предсказание вторичной структуры тРНК по алгоритму Зукера с помощью ViennaRNA представлено на рисунке 1.
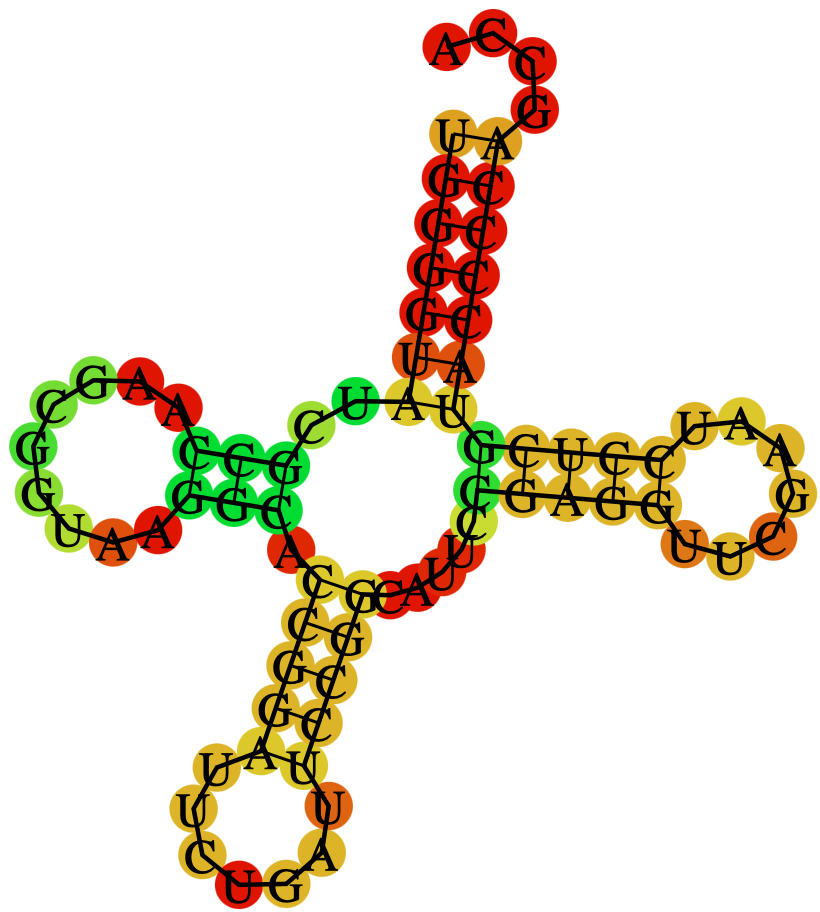 Рисунок 1. Предсказание вторичной структуры тРНК по алгоритму Зукера с помощью ViennaRNA.
Рисунок 1. Предсказание вторичной структуры тРНК по алгоритму Зукера с помощью ViennaRNA.
В таблице 1 представлено сравнение реальной и предсказанной вторичной структуры тРНК.
Таблица 1. Сравнение реальной и предсказанной вторичной структуры тРНК.
| Участок структуры |
Позиции в структуре (по результатам find_pair) |
Результаты предсказания с помощью einverted |
Результаты предсказания по алгоритму Зукера |
| Акцепторный стебель |
902-907, 966-971 |
902-907, 966-971 |
Предсказано 6/6 пар |
| D-стебель |
910-912, 923-925 |
– |
Предсказано 3/3 пар |
| T-стебель |
949-953, 961-965 |
– |
Предсказано 5/5 пар |
| Антикодоновый стебель |
937-944, 926-933 |
– |
Предсказано 5/8 пар |
| Общее число канонических пар нуклеотидов |
19 |
6 |
20 |
Задание 2
В данном задании использовалась структура 1P47.
При помощи программы define JMol были последовательно заданы множества атомов кислорода 2'-дезоксирибозы, кислорода в остатке фосфорной кислоты и азота в азотистых основаниях.
Скрипт для множеств атомов
Далее был написан скрипт, который даёт последовательное изображение всей структуры ДНК, структуры ДНК в проволочной модели и этой же структуры с выделенными шариками множествами атомов из предыдущего скрипта.
Скрипт для последовательного изображения структуры
Следующей целью было рассмотрение ДНК-белковых контактов в заданной структуре.
Я считала атомы кислорода и азота полярными, а атомы углерода, фосфора и серы – неполярными.
Полярным контактом я считала ситуацию, в которой расстояние между полярным атомом белка и полярным атомом ДНК меньше 3.5Å.
А неполярным контактом – в которой расстояние между неполярным атомом белка и неполярным атомом ДНК меньше 4.5Å.
Для заданной структуры было определено количество контактов разной природы между ДНК и цепью А белка (таблица 2).
Таблица 2. Контакты разного типа в комплексе 1P47.pdb.
| Контакты атомов белка с |
Полярные |
Неполярные |
Всего |
| остатками 2'-дезоксирибозы |
1 |
8 |
9 |
| остатками фосфорной кислоты |
11 |
10 |
21 |
| остатками азотистых оснований со стороны большой бороздки |
14 |
19 |
33 |
| остатками азотистых оснований со стороны малой бороздки |
0 |
0 |
0 |
Заметим, что в ДНК-белковых взаимодействиях преобладают неполярные связи.
Также связи с остатками азотистых оснований со стороны малой бороздки не образуются (по крайней мере в случае заданной структуры). В то же время с остатками азотистых оснований со стороны большой бороздки образуется довольно много связей.
ДНК-белковые контакты были визуализированы при помощи программы nucplot (рисунок 2).
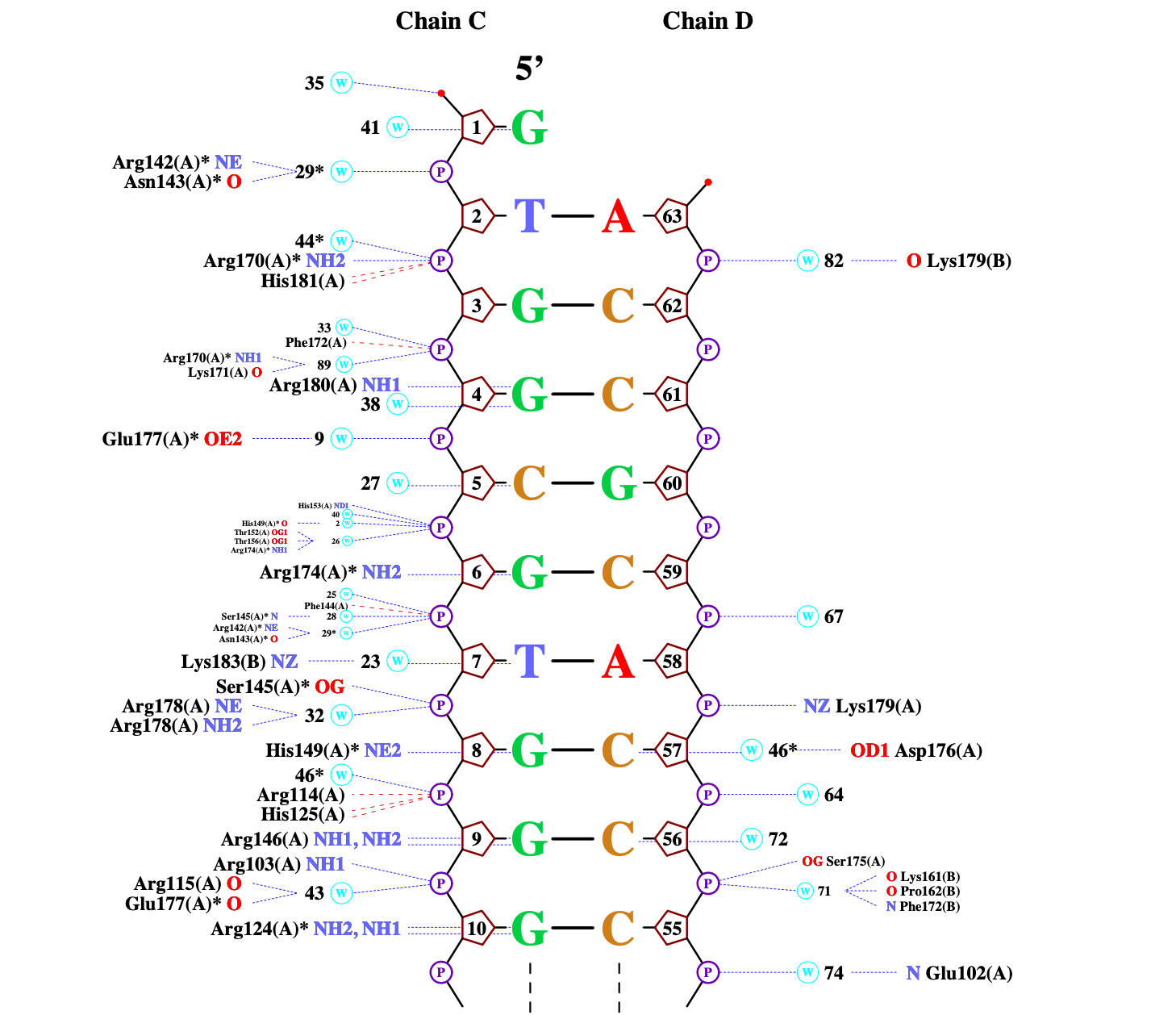
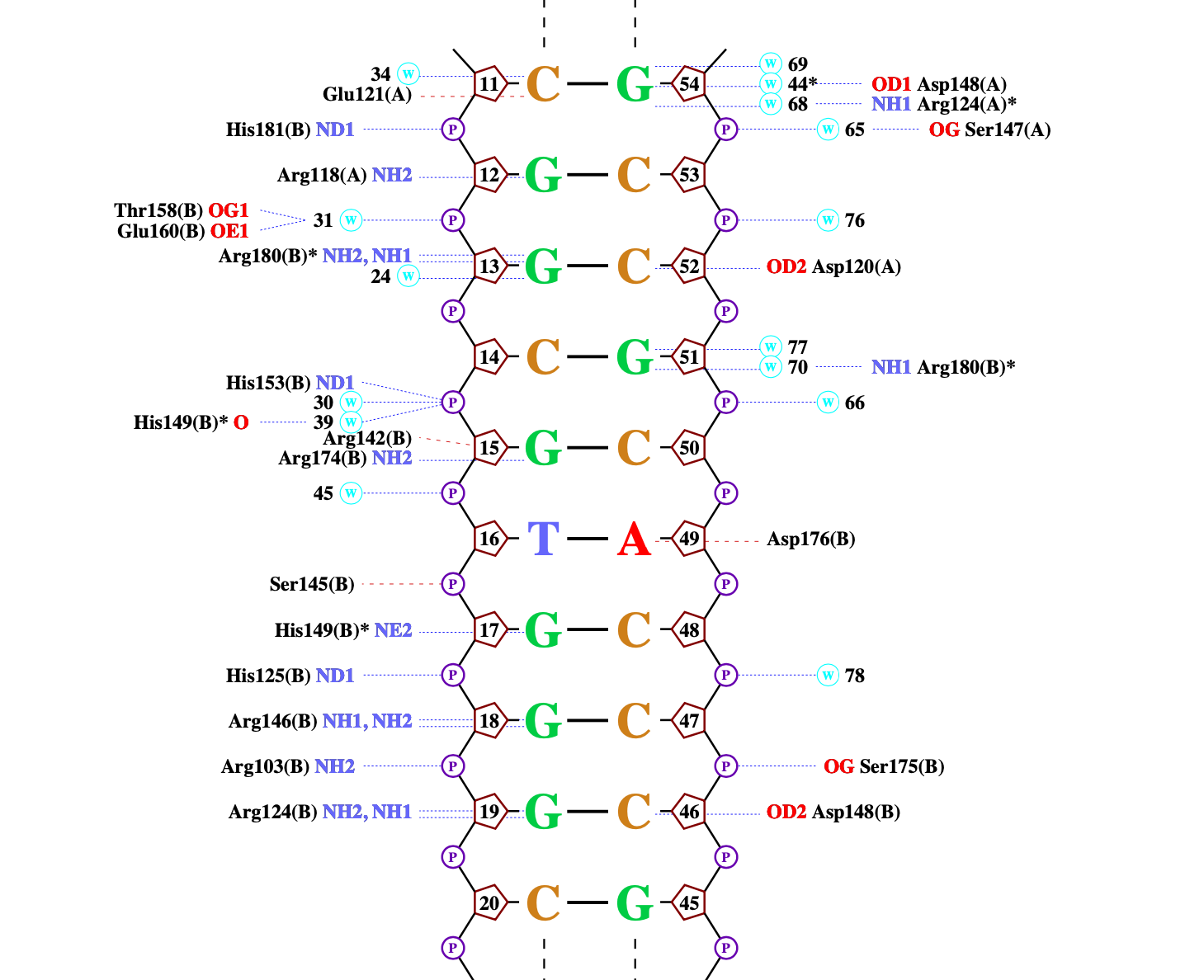
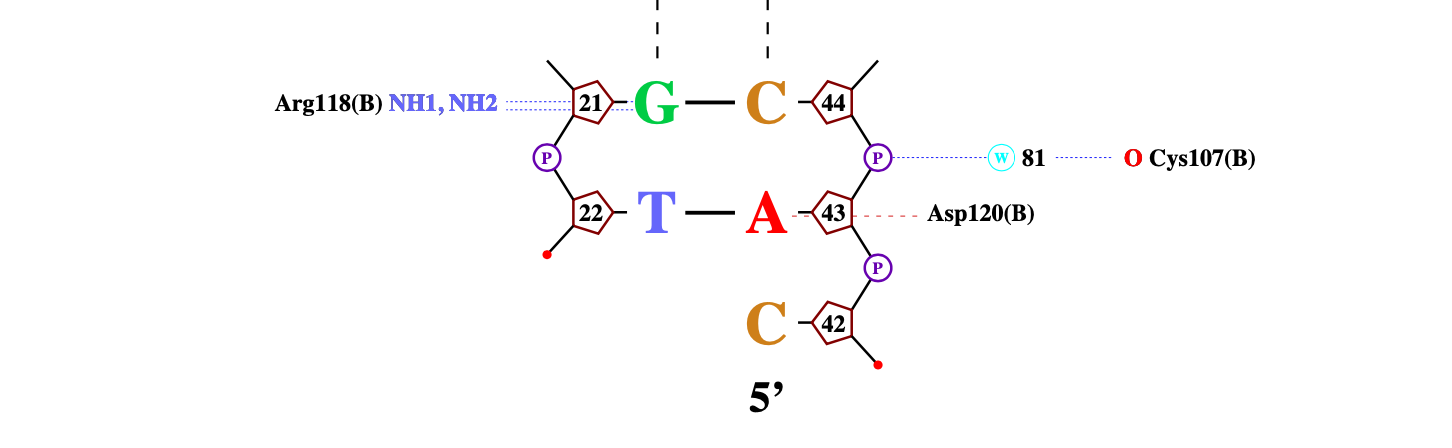
 Рисунок 2. Схема ДНК-белковых взаимодействий, полученная с помощью программы nucplot.
Рисунок 2. Схема ДНК-белковых взаимодействий, полученная с помощью программы nucplot.
На данной схеме я выбрала аминокислотный остаток с наибольшим количеством контактов с ДНК. Это Arg146, который имеет 2 связи с ДНК.
Можно предположить, что этот же остаток является наиболее важным для распознавания ДНК, так как имеет наибольшее количество контактов с ДНК.
Для данного контакта при помощи программы PyMol была создана визуализация (рисунок 3).
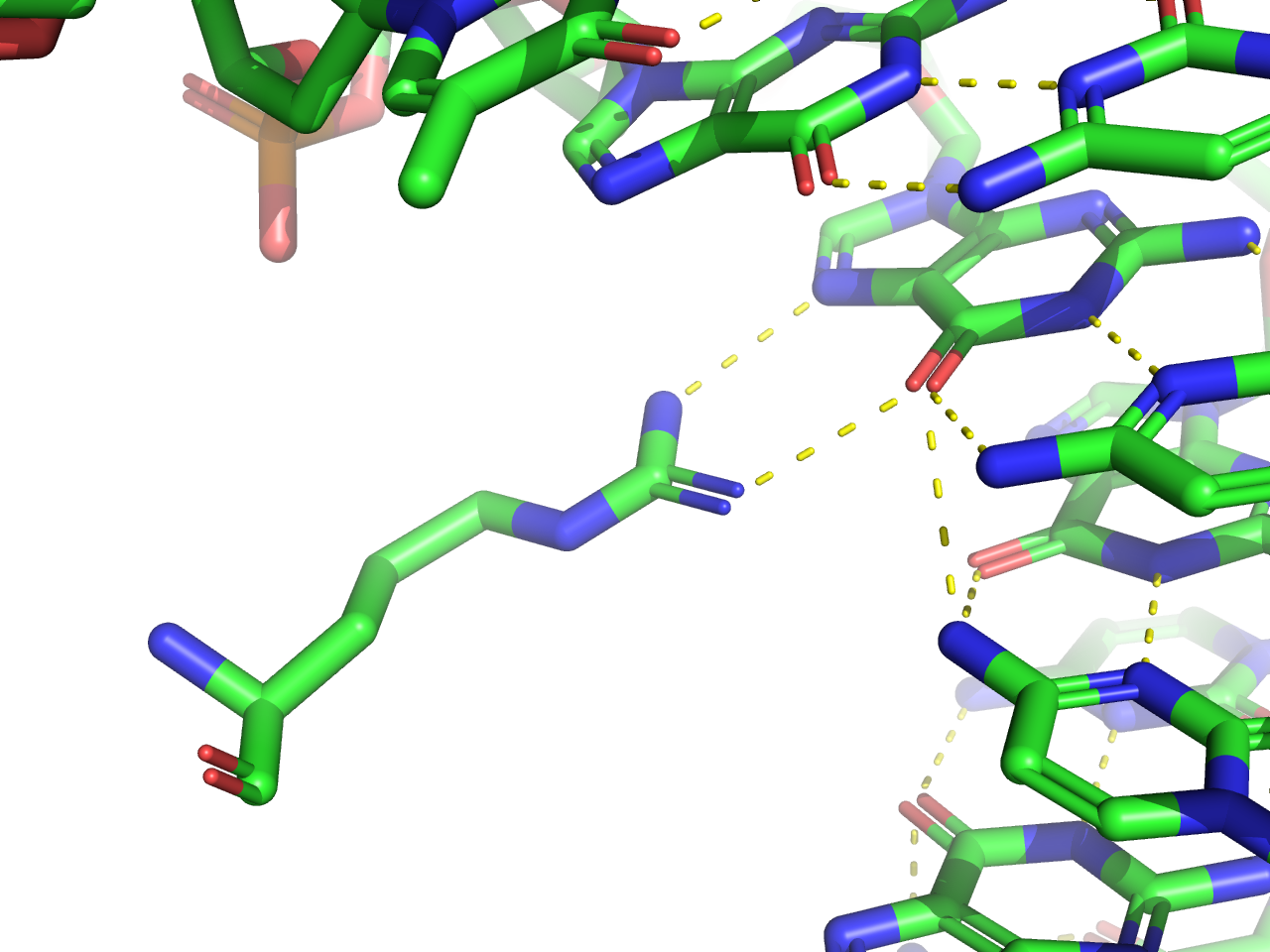 Рисунок 3. Взаимодействие Arg146 с ДНК.
Рисунок 3. Взаимодействие Arg146 с ДНК.