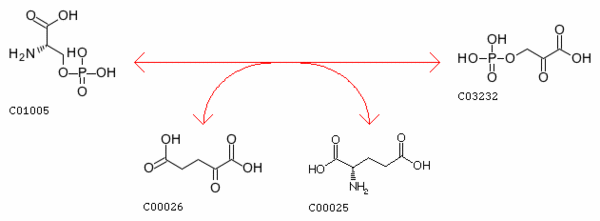
Название пути: метаболизм метионина.
Идентификаторы использованных карт: 1) eco00271 для Escherichia coli K-12 MG1655 и 2) rpr00271 для Rickettsia prowazekii.
Синтез метионина из гомосерина возможен несколькими способами: 1) через O-сукцинил-L-гомосерин (2 способа) 2) через
O-ацетил-L-гомоцистеин (2 способа) 3) через 2-оксобутаноат (1 способ).
L-метионин, кроме описанных способов из L-гомоцистеин, можно получить из N-формил-L-метионин. Гомосерин можно
получить по пути метаболизма глицина, серина и треонина из L-аспартат-4-полуальдегида.
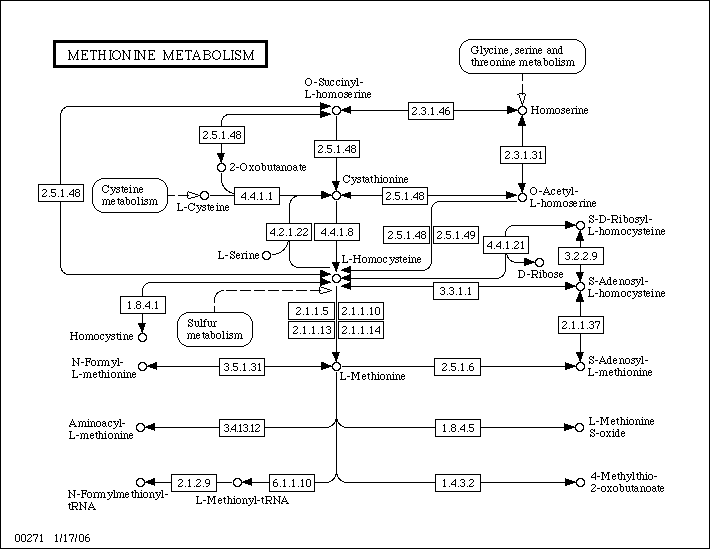
Ниже можно видеть карту заданного пути метаболизма:
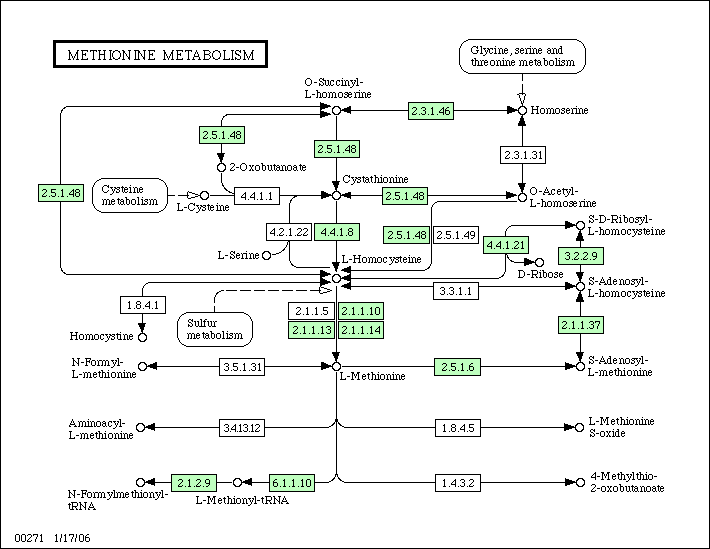
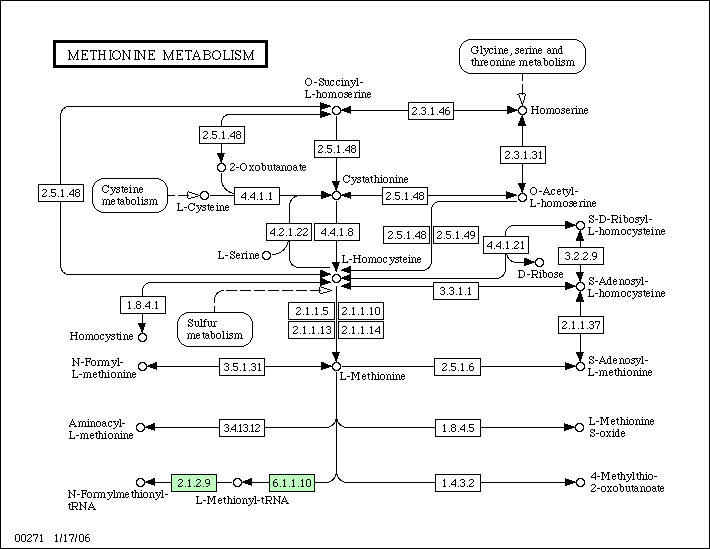
Слева можно видеть соответствующий путь для E.coli K-12, а справа - для Rickettsia prowazekii.
|
|
Предположения о биологическом смысле: видно, что E.coli K-12 имеет все необходимые ферменты для биосинтеза метионина из гомосерина
(при этом не все перечисленные раннее пути синтеза осуществимы).
Как видно, у Rickettsia prowazekii не один из возможных путей не осуществим из-за отсутствия соответствующих ферментов.
Известно, что при переходе к паразитирующему образу жизни происходит перестройка организма на новый тип работы.
Например, может редуцироваться кишечник или происходить потеря других органов. Напротив возникает большое количество
образований, которые помогают паразитировать (крючки, присоски, кутикулы и т.д.). Все эти изменения сначала происходят
на генетическом уровне. Появляются новые гены, изменяются старые. При не надобности ген становится молчащим или вовсе
может быть утрачен (зачем тратить нуклеотиды на синтез ДНК, которая никогда не пригодится). У внутриклеточного паразита
Rickettsia prowazekii при паразитировании есть все необходимое. Зачем паразиту синтезировать что-то, если вокруг этого
предостаточно. Внутри клетки метионина достаточно, его просто нужно взять, поэтому, скорей всего, развита система
гидролитических ферментов, для гидролиза белков и получения необходимых аминокислот. То есть паразит всецело зависит
от организма хозяина. Погиб хозяин - погиб паразит. Для него вещества, получаемые от хозяина, которые синтезировать
сам не в состоянии - незаменимые.
Что касается E.coli K-12, у нее нет постоянного источника аминокислот (если, конечно, организм не живет на питательной
среде; но в этом случае происходят своеобразные изменения в биохимии организма). Поэтому всегда полезно иметь возможность
синтезировать недостающее вещество из того, которое в избытке. И, самое интересное, для этого есть все необходимые
ферменты. В изменяющихся условиях среды такие организмы намного устойчивее.