Никотиновый ацетилхолиновый рецептор (АХР) является одним из первых ионных каналов, изученных в деталях. Никотиновые АХР
представлены в постсинаптических мембранах скелетных мышечных волокон позвоночных, в
нейронах нервной системы беспозвоночных и позвоночных, в синапсах электрического органа
электрического ската.
Эти рецепторы активируются ацетилхолином (АХ), освобождающимся из пресинаптических нервных
окончаний. При активации АХР открываются ионные каналы, через которые катионы могут
проходить клеточную мембрану постсинаптической клетки. Рецепторы обозначаются как
«никотиновые», поскольку действие АХ воспроизводится никотином.[1]
Физические свойства АХР рецептора
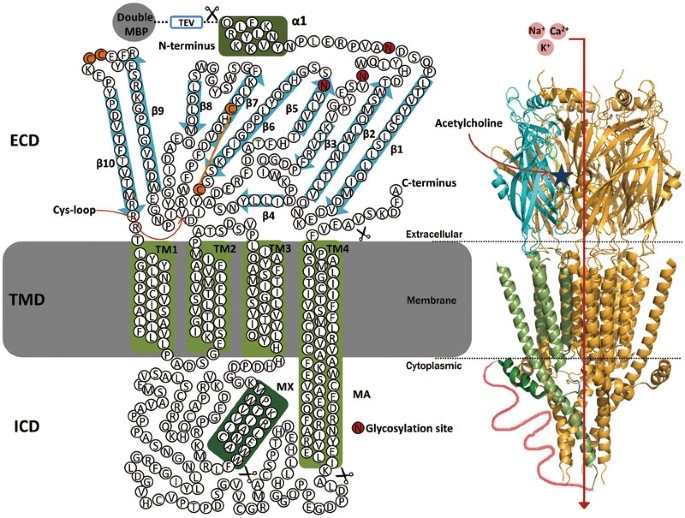
АХР представляет собой пентамер. В состав его структуры входят 5 субъединиц(две α,
одна β, одна γ и одна δ), которые образуют кольцо вокруг центральной поры.[1]
Функции канала
Семейство никотиновых рецепторов состоит из множества различных подтипов, каждый из которых имеет свои физиологические функции и биофизические свойства. Подтип никотиновых рецепторов α3β4 образует главный переключатель между центральной и периферической нервными системами в вегетативных ганглиях. Этот рецептор также локально экспрессируется в областях мозга, которые отвечают за систему вознаграждения и зависимость.
Вегетативная нервная система включает симпатические и парасимпатические пути и осуществляет любой непроизвольный контроль над внутренними органами. Симпатическая ветвь, как правило, формирует ответ на острый стресс по типу "бей или беги", тогда как парасимпатическая - поддерживает периферический гомеостаз. Несмотря на их антагонистические эффекты, обе ветви обычно сохраняют дисинаптическую архитектуру. Нейроны в вегетативных ганглиях, которые иннервируют нижестоящие мишени, получают холинергический сигнал от центральной нервной системы. Преобладающим рецептором синаптической передачи в этих нейронах является никотиновый ацетилхолиновый рецептор α3β4, обычно называемый ганглионарным никотиновым рецептором.[2]
Хотя этот рецептор является важным в вегетативной нервной системе, его экспрессия этим ограничивается. Рецептор α3β4 также в изобилии обнаружен в хабенуло-межпеданкулярном тракте, который модулирует мезолимбическую дофаминовую систему, основной путь вознаграждения в мозге. На линиях грызунов было показано, что функциональные антагонисты α3β4, такие как 18-метоксикоронаридин и AT-1001, уменьшают самостоятельное потребление широкого спектра наркотических веществ, включая морфин, кокаин, алкоголь и никотин. Полученные данные вызвали значительный интерес использования данных подтипов в качестве мишени для терапевтических средств против зависимости.[2]
Важные аминокислотные последовательности.
В первичной последовательности белка обнаружены обширные участки
неполярных аминокислот, способных к образованию
трансмембранного домена. В оригинальной модели АХР были идентифицированы четыре такие области (TМ1-TМ4, см. рис. 1.).
На NН2-конце находится относительно гидрофобный участок из 24
аминокислот. Эта последовательность является сигнальной. Она необходима для
проникновения синтезированного в клетке белка АХР в поверхностную мембрану.
Следовательно, NН2-конец является внеклеточным.[1]
Общая информация о белке
PDB id: 6PV7
Uniprot id: ACHB4_HUMAN
Название белка: Human alpha3beta4 nicotinic acetylcholine receptor in complex with nicotine
Молекулярная масса: 411.62 кДа
Число аминокислот: 2379
Описание лигандов
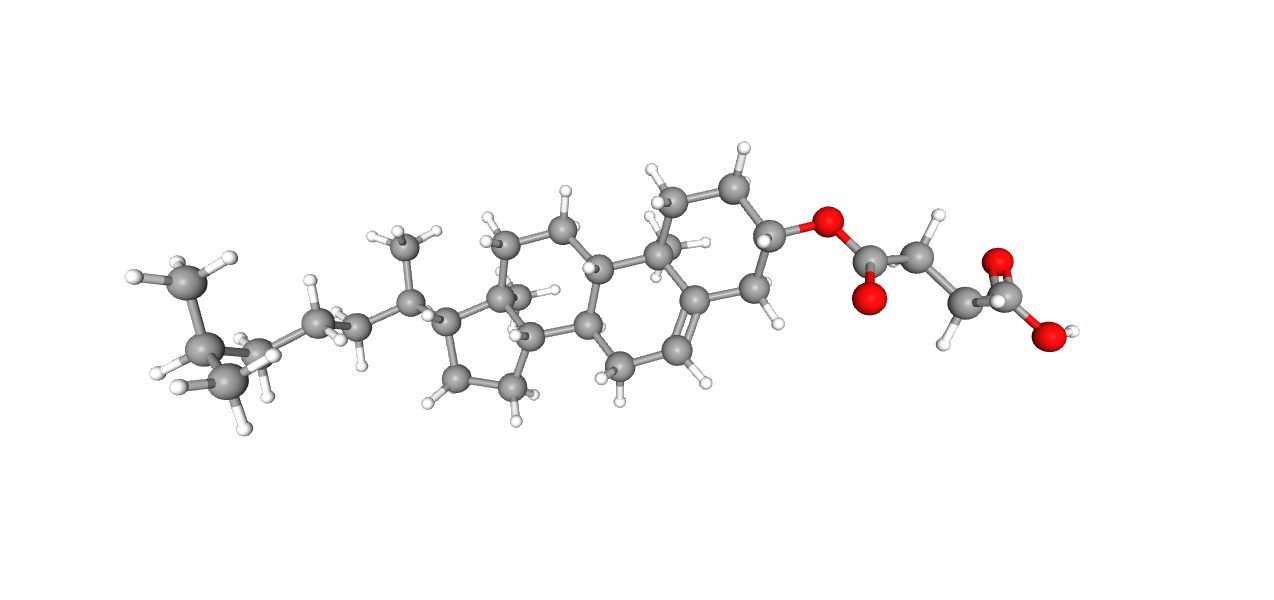
Холестерола гемисукцинат
Название по IUPAC: 4-[[(3S,8S,9S,10R,13R,14S,17R)-10,13-dimethyl-17-[(2R)-6-methylheptan-2-yl]-2,3,4,7,8,9,11,12,14,15,16,17-dodecahydro-1H-cyclopenta[a]phenanthren-3-yl]oxy]-4-oxobutanoic acid
Брутто-формула: C31H50O4
Молярная масса: 486.7
Ссылка на идентификатор в базе данных PubChem: 65082
Ацетилглюкозамин
Название по IUPAC: 2-acetamido-2-deoxy-beta-D-glucopyranose
Брутто-формула: C8H15NO6
Молярная масса: 221.21
Ссылка на идентификатор в базе данных PubChem: 24139
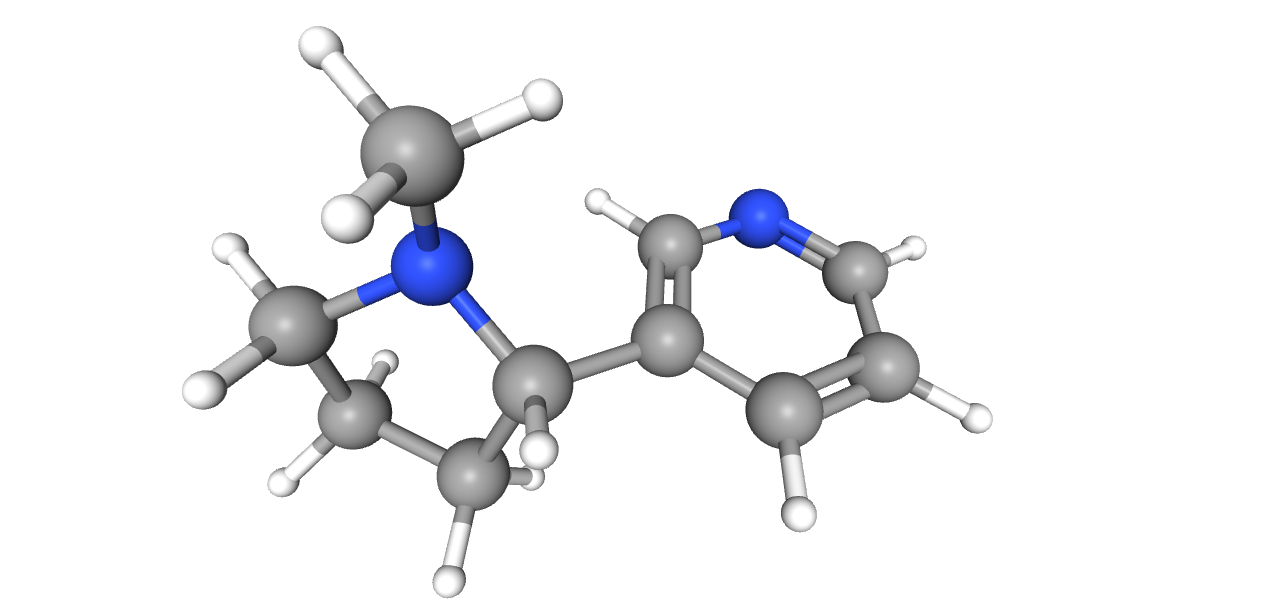
Никотин
Название по IUPAC: 3-[(2S)-1-methylpyrrolidin-2-yl]pyridine
Брутто-формула: C10H14N2
Молярная масса: 162.23
Ссылка на идентификатор в базе данных PubChem: 89594
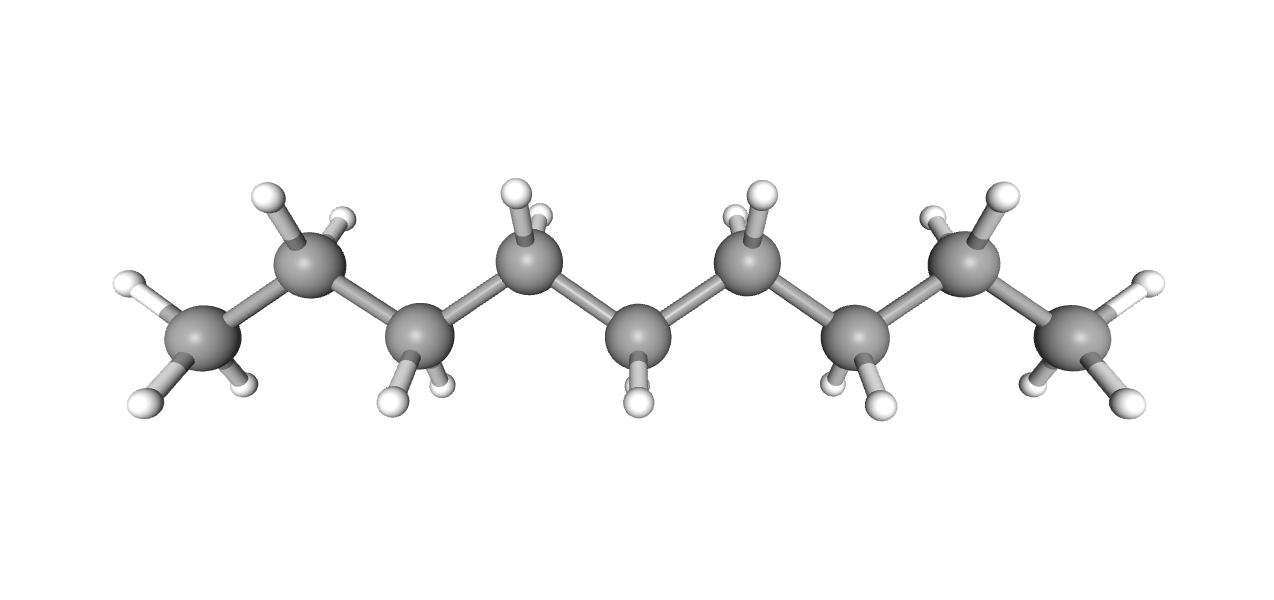
Нонан
Название по IUPAC: nonane
Брутто-формула: C9H20
Молярная масса: 128.25
Ссылка на идентификатор в базе данных PubChem: 8141
Ион натрия
Название по IUPAC: sodium ion
Брутто-формула: Na
Молярная масса: 22.99
Ссылка на идентификатор в базе данных PubChem: 923
Взаимодействия белок-лиганд и их роль в структуре
Холестерола гемисукцинат
Белок анализировался в липидных нанодисках, содержащих, помимо прочего, холестеролгемисукцинат - производное холестерина, близкое к нему функционально.[2] Как и ожидалось, исходя из природы и функции соединения, оно связано с трансмембранными альфа-спиралями белка гидрофобными взаимодействиями - к примеру, с остатками Ile282:A, Leu286:A, Val289:A, Ile 290:A и Phe293:A.
Ацетилглюкозамин
N-ацетил-гликозамины, указанные на странице pdb как лиганды, яаляются на самом деле модификациями остатков аспарагинов, для которых не была указана связь. Все моно- или олигосахариды расположены у внеклеточной части белка. Для каждого из них на расстоянии не более 1,5 ангстрем от атома углерода находится ровно один "чужой" атом -азот аспарагина (см. скрипт), что позволяет предположить наличие между ними ковалентной связи там, где она не показана. Наконец, общее количество и расположение моно- и олигосахаридов - 16 - соответствует количеству и расположению гликозилированных аспарагинов для двух альфа- (A, D; по 2 остатка)[3] и трёх бета-цепей (B, C, E; по 4 остатка)[4] по данным UniProt.
Никотин
Никотин связан пи-катионным стекингом с пятью аминокислотными остатками на петлях A-D (для сайта связывания, образованного цепями D и E: A: Tyr93:D, B: Trp149:D, C: Tyr190:D, Tyr197:D, D: Trp59:E). Особенно важным является триптофан B-петли, с карбонильной группой которого никотин также связан водородной связью.[5] Атом воды, удалённый от незаряженного азота никотина на 3 ангстрема и от кислорода остова Asn111 на 2.4 ангстрема с углом 139 градусов, вероятно, образует мостик между названными атомами.
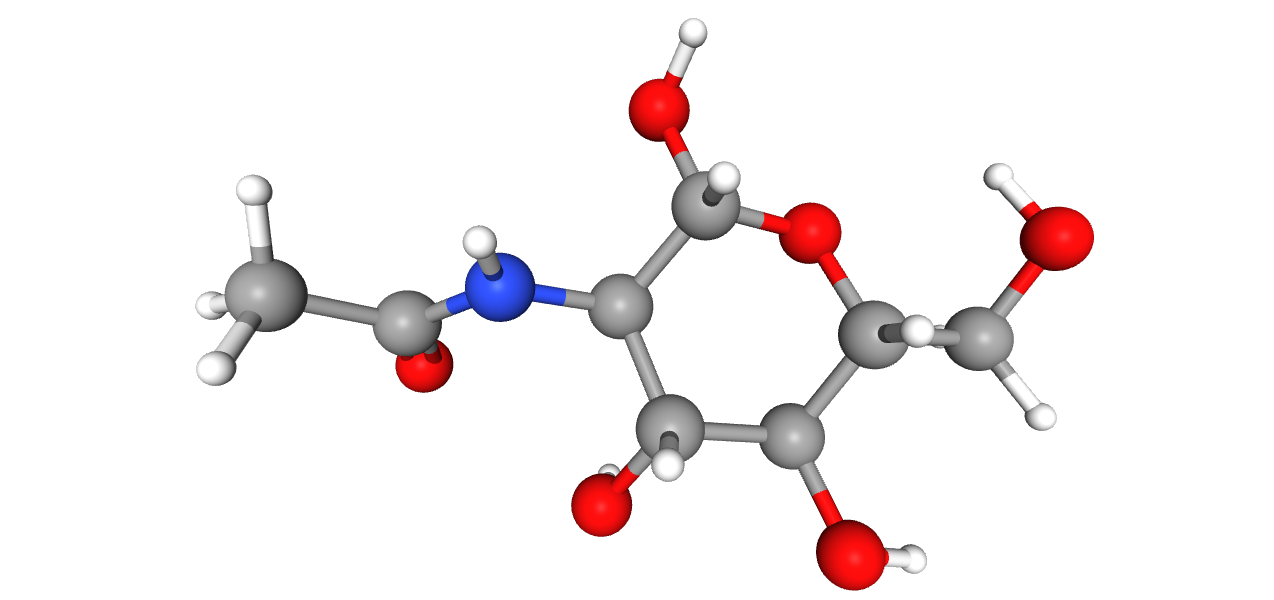
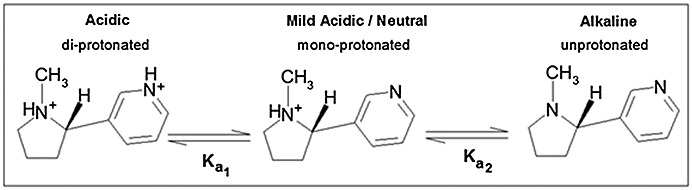
Схема 1. Протонирование никотина
По табличным значениям констант кислотности функциональных групп в составе никотина можно сделать вывод, что в клетке эта молекула находится в моно-протонированном состоянии (схема 1).
Нонан
Нонан отражает хвост молекулы детергента или липида, занимающий, судя по картине электронных плотностей, наиболее удалённую от мембраны часть канала во внутриклеточном домене.
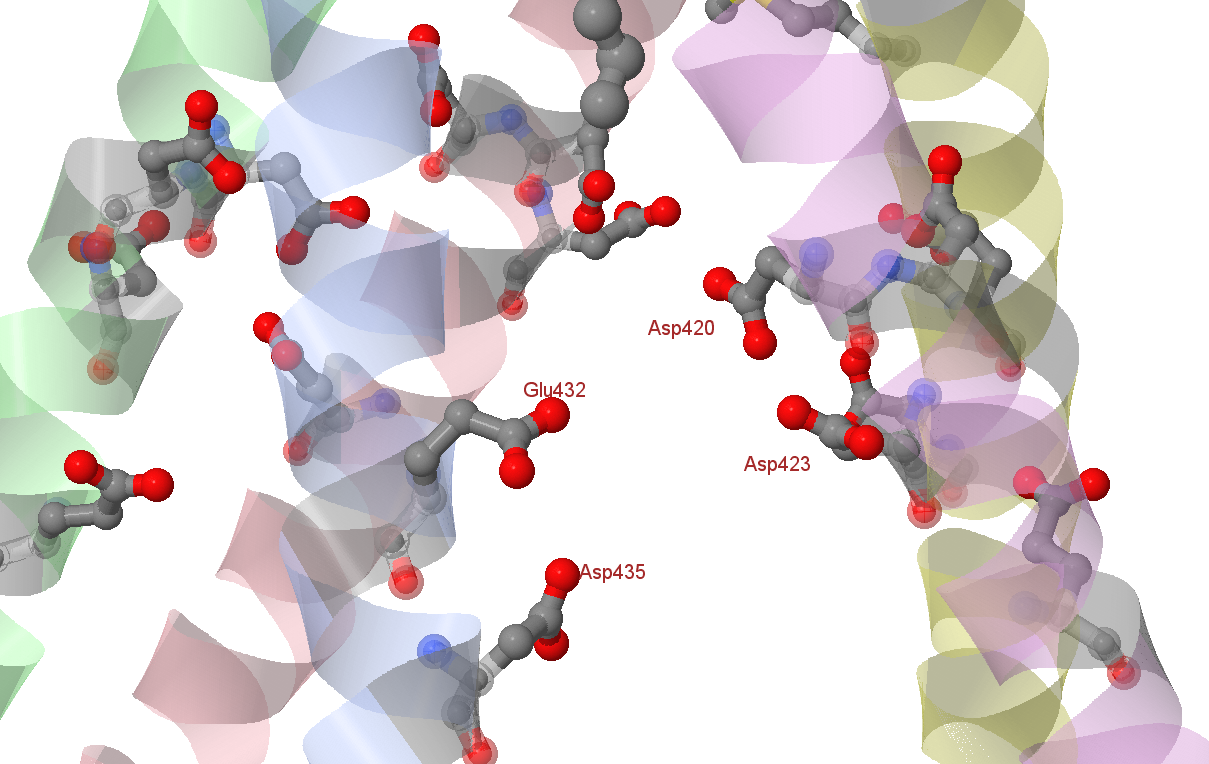
Гидрофобные остатки альфа-спиралей в этом регионе, такие как Val417, Ile420, и Met424 альфа-цепей и Val410, Ile413 и Met417 бета-цепей, консервативны; данные молекулярной динамики говорят о том, что липид должен занимать этот сайт для поддержания стабильности внутриклеточного домена. Всё это говорит в пользу того, что данный регион может заниматься гидрофобной молекулой in vivo и подтверждает существующий консенсус о том, что ионы покидают канал не через данную гидрофобную "пробку" в его конце, а через пять боковых "окон" внутриклеточного домена, содержащих остатки аспартата и глутамата (в α-цепях - Glu432 и Asp435, в β-цепях - Asp420, Asp421, Asp423 и Glu428), замена которых на лизины значительно снизила проводимость канала (рис. 2).[2] Эти остатки также продемонстрированы в скрипте.
Рис. 2. Боковые "окна" внутриклеточного домена, через которые Na+ покидает канал с гидрофобной "пробкой". Внутриклеточная среда на картинке сверху.
Ион натрия
Ион натрия находится в ближней к цитоплазме половине трансмембранного домена, вблизи самого узкого места канала, образованного остатками глутамата (Glu240 в альфа- и Glu241 в бета-цепях), электростатически взаимодействующими с металлом. Из схожести исследуемой структуры белка со структурой, полученной в присутствии детергентов, и из того факта, что вся трансмембранная область канала сольватирована, исследователями был сделан вывод, что данная структура отражает канал в непроводящем десенситизированном состоянии. При этом предполагается, что остатки глутаматов, ориентирусь вовнутрь, образуют сайт связывания катиона.[2]Остатки треонина (Thr243 в альфа- и Thr244 в бета-цепях) в непосредственной близости к иону, возможно, в проводящем состоянии играют роль фильтра, избирательно пропускающего натрий.
Информация о белок-белковых контактах
Субъединицы белка не связаны между собой ковалентно. Однако в нем присутствуют немногочисленные солевые мостики (разновидность ионной связи между COO- и NH3+ группами). Практически все обнаруженные водородные связи расположены внутри альфа спиралей и бета листов, так как они играют неотъемлемую роль в формировании вторичной структуры белка. Кроме того, была выявлена внутрисубъединичная связь между петлями B и C, которая обеспечивает высокое сродство данного типа рецепторов к нейромедиаторам.[2]
Основной вклад в поддержании третичной структуры вносят гидрофобные взаимодействия и стэкинг.
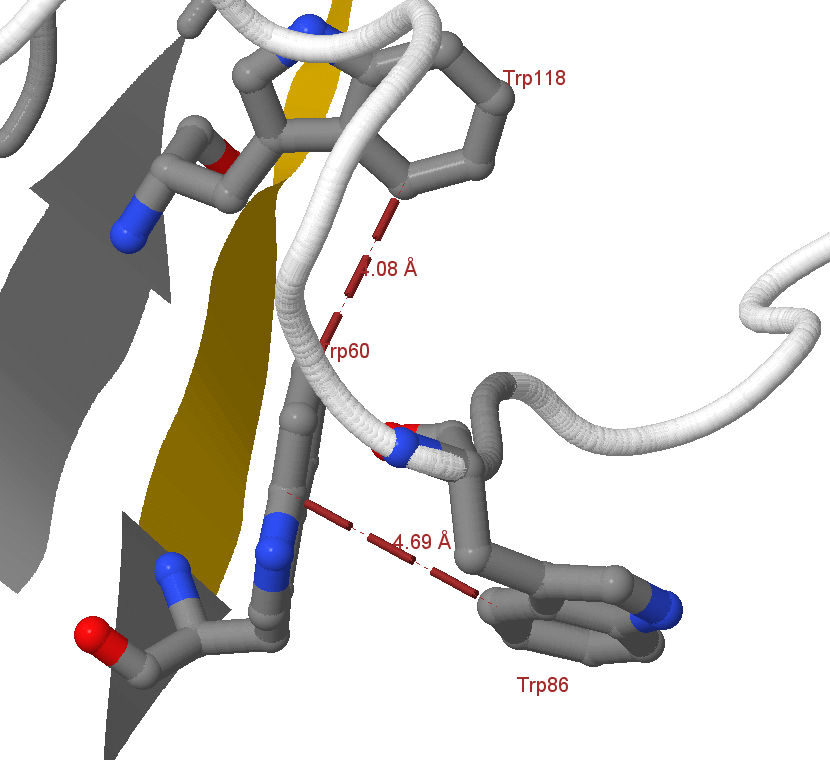
Рис.3. Т-стэкинг
Стэкинг
Для поиска стекинг-взаимодействий внутри белка был использован портал PIC. Результат выдачи вы можете посмотреть по ссылке.
Взаимодействия между ароматическими остатками.
Формально, данные взаимодействия определяются как пара остатков ароматических аминокислот, центры которых расположены на расстоянии от 4.5 до 7 Å, а двугранный угол между плоскостями ароматических групп составляет 30-90°.[6]
Для иллюстрации примера Т-стэкинга были выбраны три остатка триптофанов цепи D - W60, W86 и W118. Триптофан 60, в данном случае, может одновременно взаимодействовать с оставшимися, что изображено на рисунке 3. Данные взаимодействия удовлетворяют критериям, описанным выше (параметры приведены в таблице ниже), так что со структурной точки зрения их можно считать Т-стэкингом.
Остаток 1
Остаток 2
Расстояние между ароматическими центрами (Å)
Двугранный угол между плоскостями ароматических колец (°)
Trp60:A
Trp86:A
6.41
93.75
Trp60:A
Trp118:A
6.55
82.71
π-катионный стекинг
Другим типом стекинга, рассмотренным в работе, является полярный π-катионный стекинг. Этот тип взаимодействий имеет электростатическую природу и проявляется при взаимодействии π-электронного облака ароматического кольца с катионами. Наиболее правдоподобным критерием существования данного типа взаимодействий служит энергия образующейся связи,[7] однако для предсказаний было использовано пороговое значение в 6Å и некомпланарное расположение положительно заряженной группы с углеродами ароматического кольца.
В составе белка было рассмотрено взаимодействие Arg6:A с Tyr72:A. Расстояние между гуанидиновой группировкой и центром ароматического кольца составляло 5 Å, а двугранный угол - 118° (по результатам выдачи PIC). Хоть это и не хрестоматийный пример, однако положительный заряд аргинина способен взаимодействовать с электронным облаком на таком расстоянии, так как смещен относительно положительно заряженной плоскости ароматического кольца в сторону электронного облака.
Взаимодействия между ароматичными остатками или π-катионные взаимодействия между различными цепями белка отсутствуют.
Гидрофобное ядро
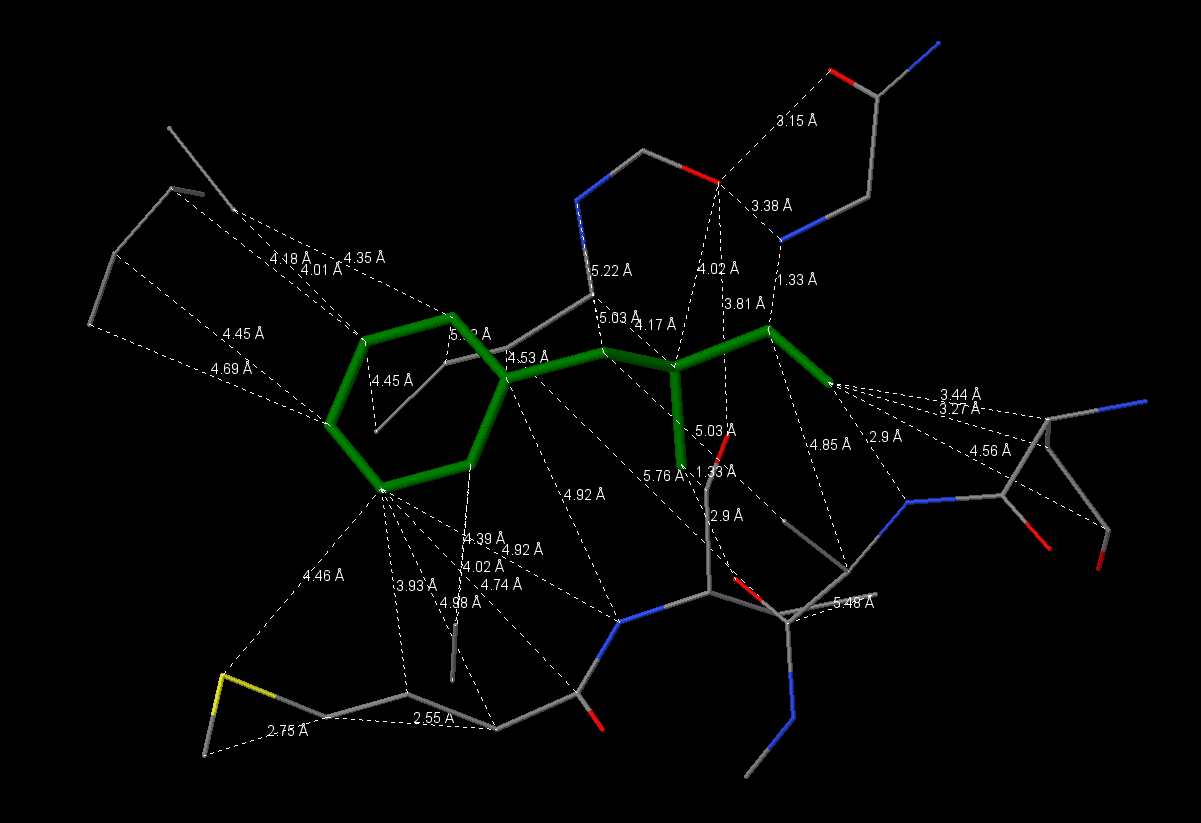
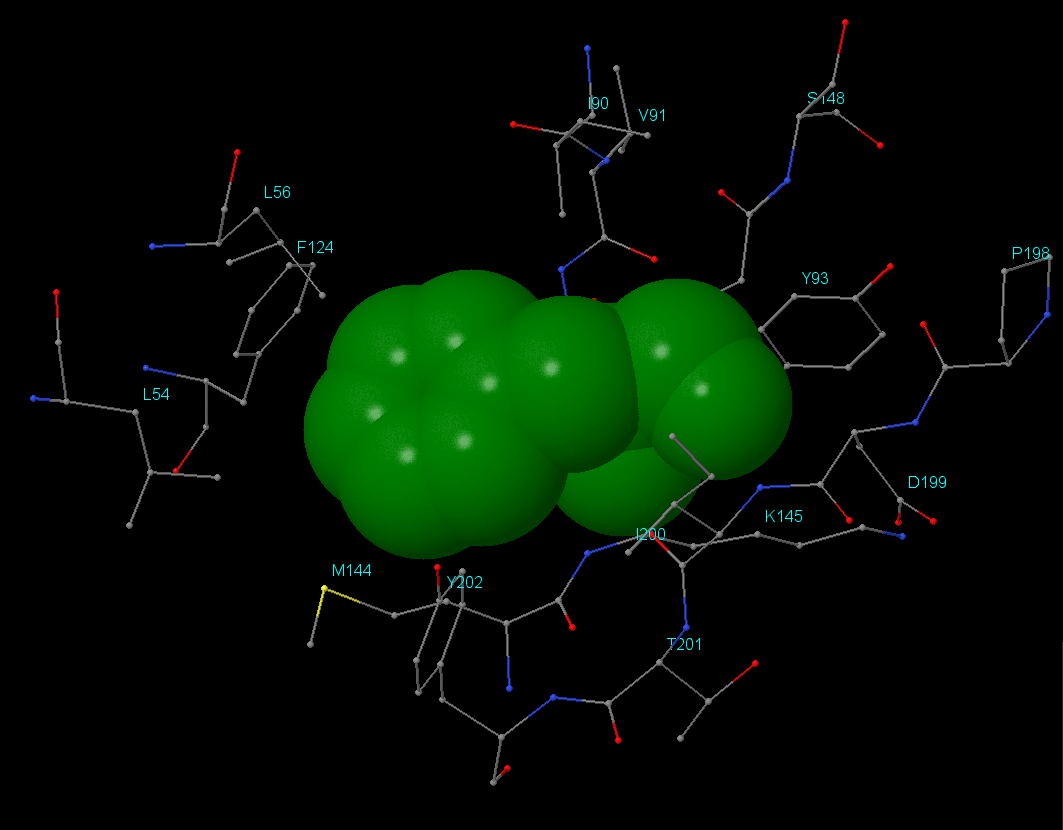
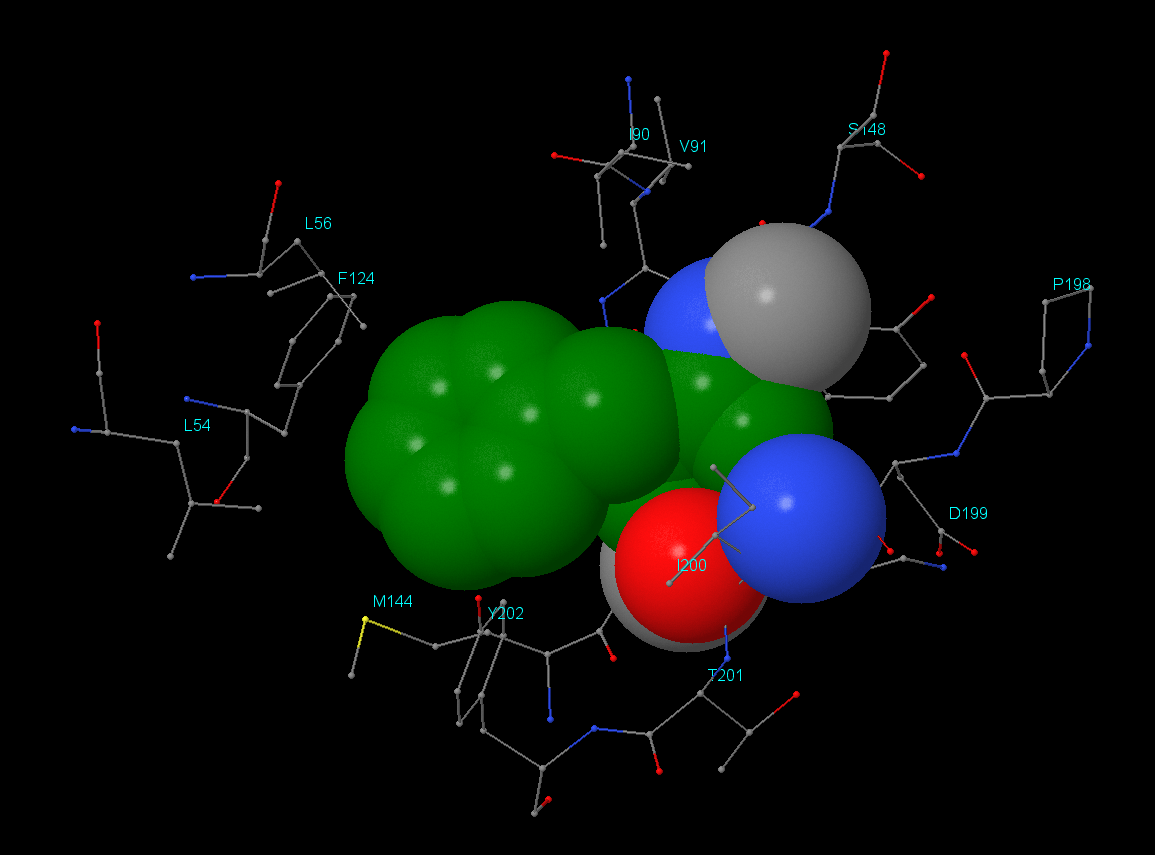
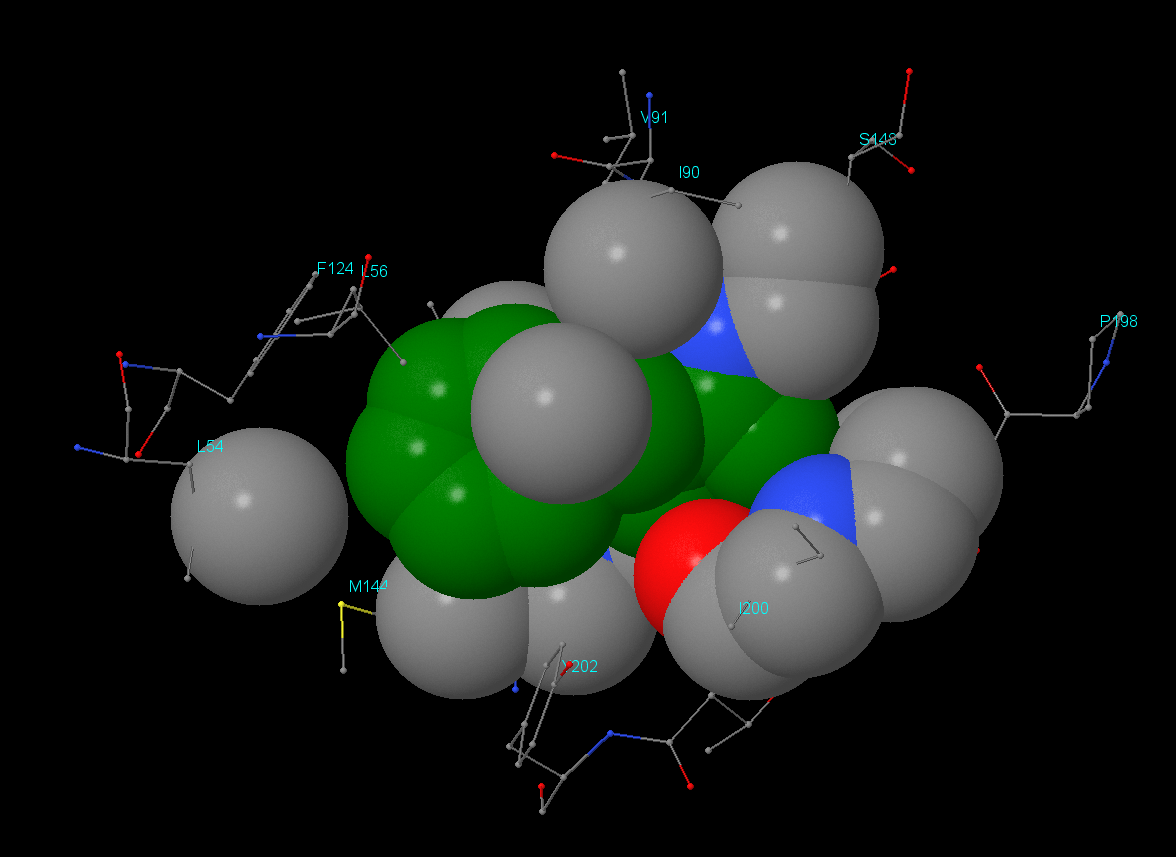
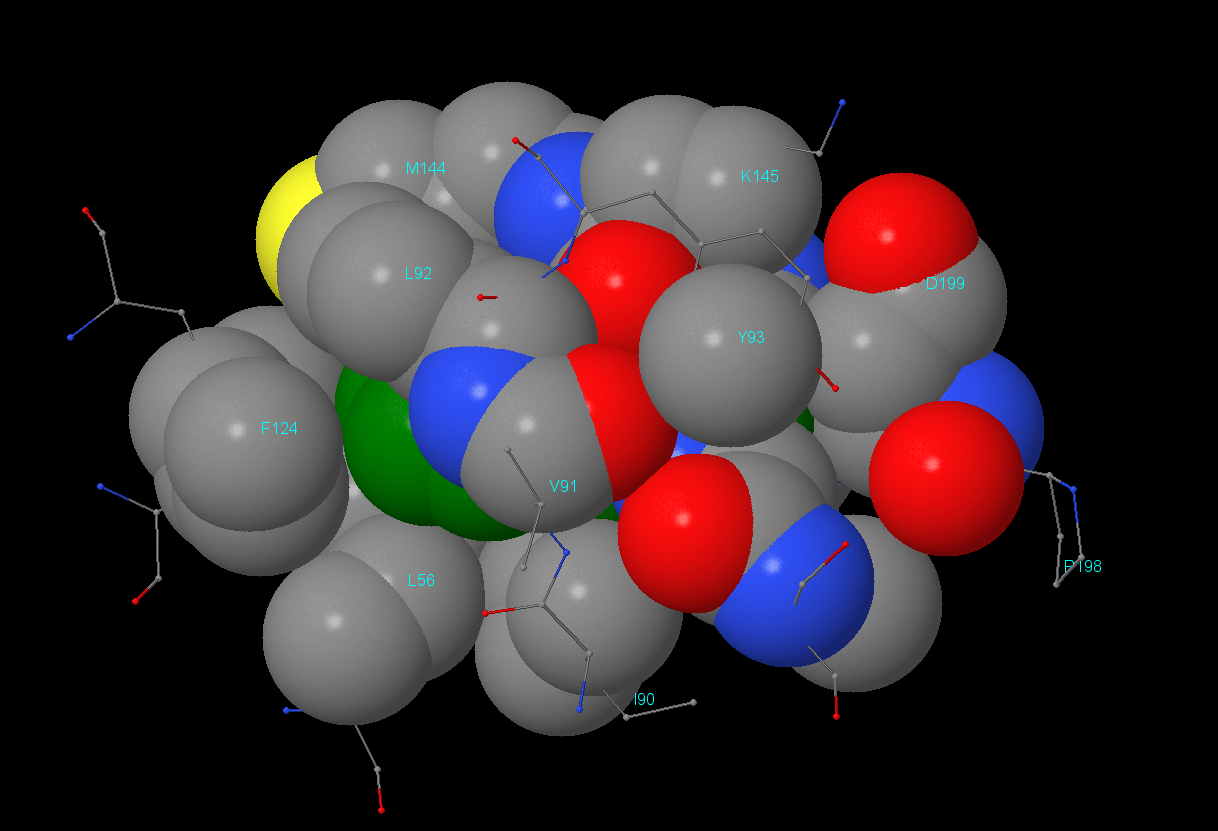
Для поиска гидрофобного кора был взят остаток Phe146 цепи А. Результат анализа можно посмотреть в JSMol-aпплете. Было выяснено, что в окружении данного остатка находятся как гидрофильные, так и гидрофобные аминокислоты, но, что интересно, контакт с Phe146 осуществляют только гидрофобные части окружающих остатков. Например, Lys145 своей ε-аминогруппой направлен от Phe146, аналогично Asp199 - его СООН-группа находится на удалении. Значительную часть гидрофобного кора составляют гидрофобные аминокислоты (алифатические(Ley, Ile, Val) и ароматические(Phe, Tyr)).
Плотность упаковки: на расстоянии 1Å атомы отсутствуют, а начиная с 2Å уже присутствуют. Остаток Phe146 практически полностью экранируется начиная с расстояния 5-6Å.
По измерению расстояний между атомами в гидрофобном ядре было получено среднее значение 4,19Å и стандартное отклонение 1,02Å (ссылка на pdf-файл).По рассчетам растояний с учетом Ван-дер-ваальсовых радиусов атомов (ссылка на электронную таблицу) было установлено, что в некоторых случаях молекула воды может располагаться в промежутках ядра.
Примечание:
Дополнительно в программе IBM SPSS Statistics был проведен статистический анализ (одновыборочный K-S-тест), показавший, что эмпирическое распределение не соответствует теоретическому (нормальному или равномерному), а значит, для него, в первом приближении, нельзя подобрать соответствующую теоретической функцию распределения и предсказывать по ней иные случаи.
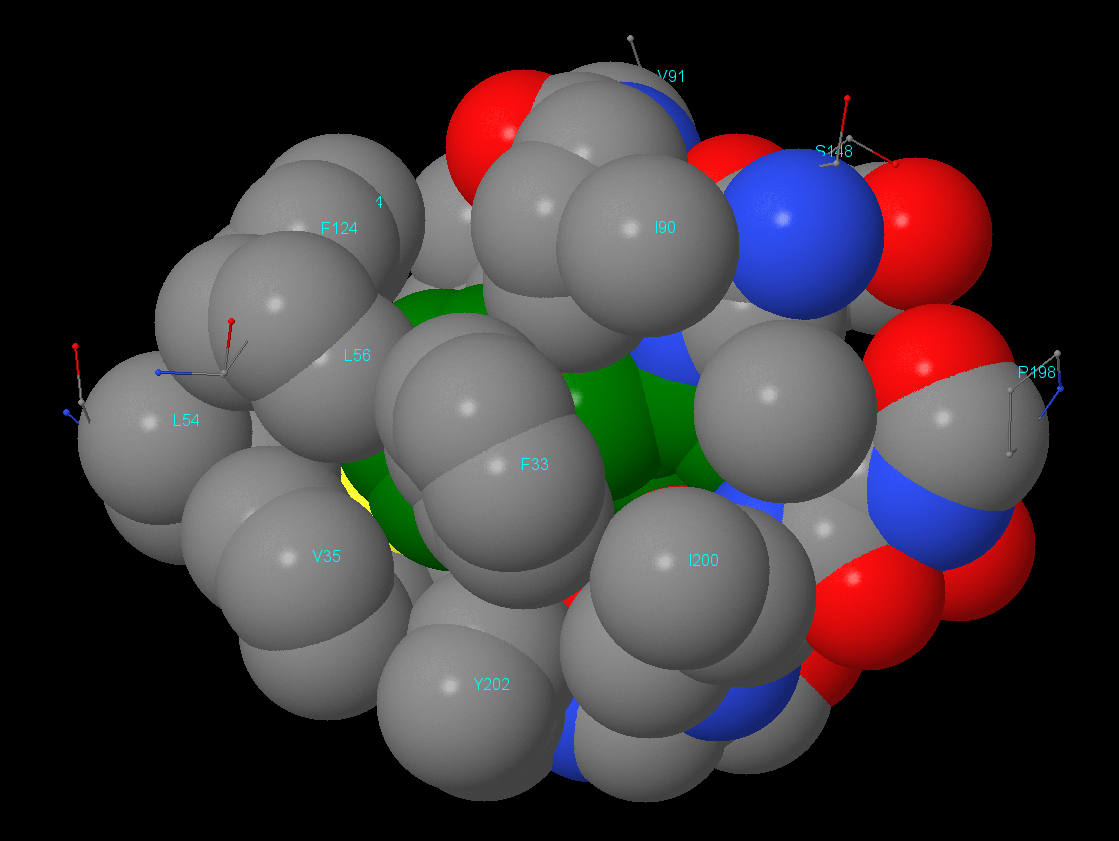
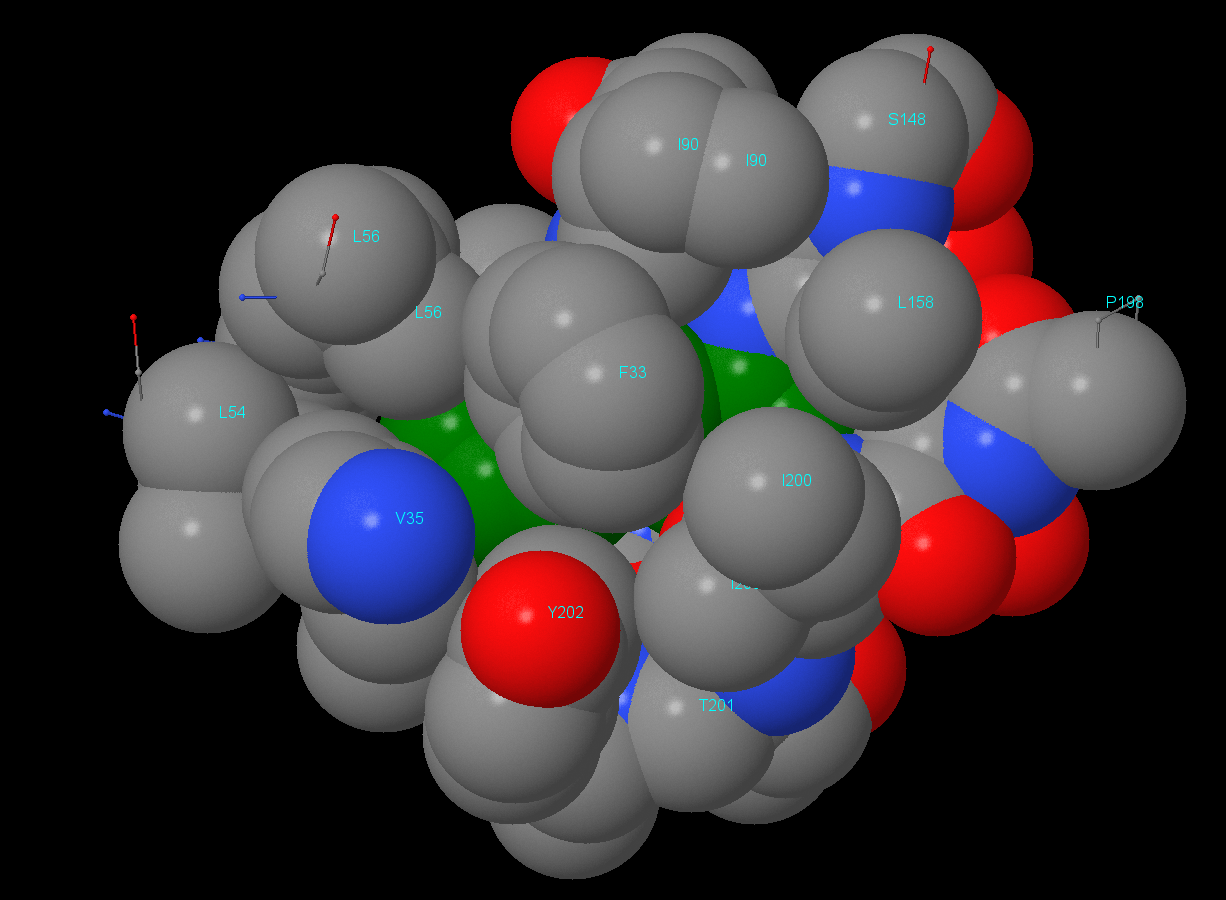
Изображения гидрофобного ядра. В данном разделе отсутствует изображение атомов в радиусе 2 Å. Остальные изображения идут в порядке увеличения расстояния от остатка Phe146 с 3 до 7 Å (шаг в 1 Å).
Бетенькова Рената написала скрипты на отображение лигандов, собирала про них общую информацию, а также исследовала дисульфидные связи в белке и написала соответствующий скрипт.
Кузьменко Олег исследовал упаковку атомов в гидрофобном ядре, изучил стекинг-взаимодействия в белке, написал соответствующие скрипты, а также координировал работу команды и помогал с оформлением веб-страницы.
Машкова София занималась оформлением отчета на веб-странице, написала введение и исследовала водородные связи и солевые мостики в белке, написав соответствующие скрипты.
Николаев Николай занимался разделом о взаимодействии лигандов с белком, написал соответствующие скрипты и изучил роль этих контактов в реализации функции белка.
Ссылки на источники
[1] Nicholls J. G. et al. From neuron to brain. – Sunderland, MA : Sinauer Associates, 2001. – Т. 271.