Задание 1. Предсказание вторичной структуры заданной тРНК
Упражнение 1. Предсказание вторичной структуры тРНК путем поиска инвертированных повторов
1eiy.fp - результат работы find_pair, запуск "~/term3/block1/pr2$ find_pair -t 1eiy_old.pdb 1eiy.fp"| Участок структуры | Позиции в структуре (по результатам find_pair) | Результаты предсказания с помощью einverted | Результаты предсказания по алгоритму Зукера |
|---|---|---|---|
| Акцепторный стебель | 5'-01-07-3' 5'-66-72-3' Всего 7 пар |
Предсказано 7 пар | Предсказано 8 пар - дополнительная 8U-G65 |
| D-стебель | 5'-10-13-3' 5'-22-25-3' Всего 4 пары |
0 пар | Все 4 пары |
| T-стебель | 5'-49-52-3' 5'-62-65-3' Всего 4 пары |
0 пар | Все 4 пары |
| Антикодоновый стебель | 5'-26-31-3' 5'-39-44-3' Всего 6 пар |
5 пар | 5 пар |
| Общее число канонических пар нуклеотидов | 22 | 24, верных 12 | 21 |
Предсказание вторичной структуры программой einverted:
Параметры:
Gap penalty [12]: 0 - потому, что у нас нет ограничений на петли.
Minimum score threshold [50]: 0 - у нас 1 структура, поэтому пусть выдает всё, что найдется
Match score [3]: 20
Mismatch score [-4]: -4
Насколько я понимаю, программа "складывает структуру поплам", поэтому не удивительно, что боковые петли она предсказывать не может
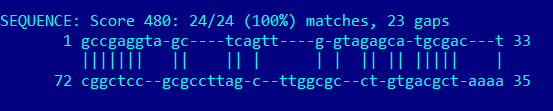
Предсказание вторичной структуры программой RNAfold:
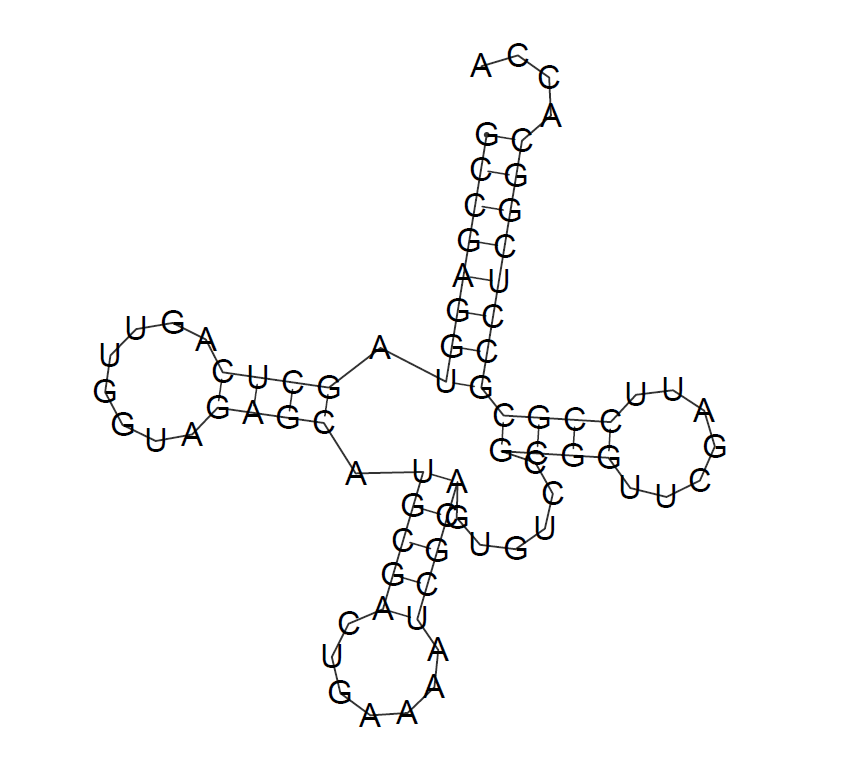 Вышло очень даже неплохо
Вышло очень даже неплохо
Задание 2. Поиск ДНК-белковых контактов в заданной структуре
Упражнение 1.
Упражнение 2.
Примеры использованных команды смотреть в файле
| Контакты атомов белка с | Полярные | Неполярные | Всего |
|---|---|---|---|
| остатками 2'-дезоксирибозы | 7 | 38 | 45 |
| остатками фосфорной кислоты | 12 | 26 | 38 |
| остатками азотистых оснований со стороны большой бороздки | 11 | 13 | 24 |
| остатками азотистых оснований со стороны малой бороздки | 3 | 4 | 7 |
Из таблицы можно заметить несколько закономерностей.
Во-первых, что сахарынй остов образует в основном неполярные
контакты.
Во-вторых, что взаимодействий в большой бороздке значительно больше, чем в малой. Скорее
всего, это объясняется бОльшими линейными размерами, а также бОльшей эффективностью взаимодействия с более
глубокой большой бороздкой.
Также видно, что с остовом в целом взаимодействий больше, чем с внутренними атомами
бороздок. Это тоже логично, тк стерически остов более доступен для подхода других молекул.
Задание 3. Nucplot: ДНК-белковые контакты
Программа Nucplot (запуск - nucplot 1hw2_old.pdb) выдала следующую картинку:
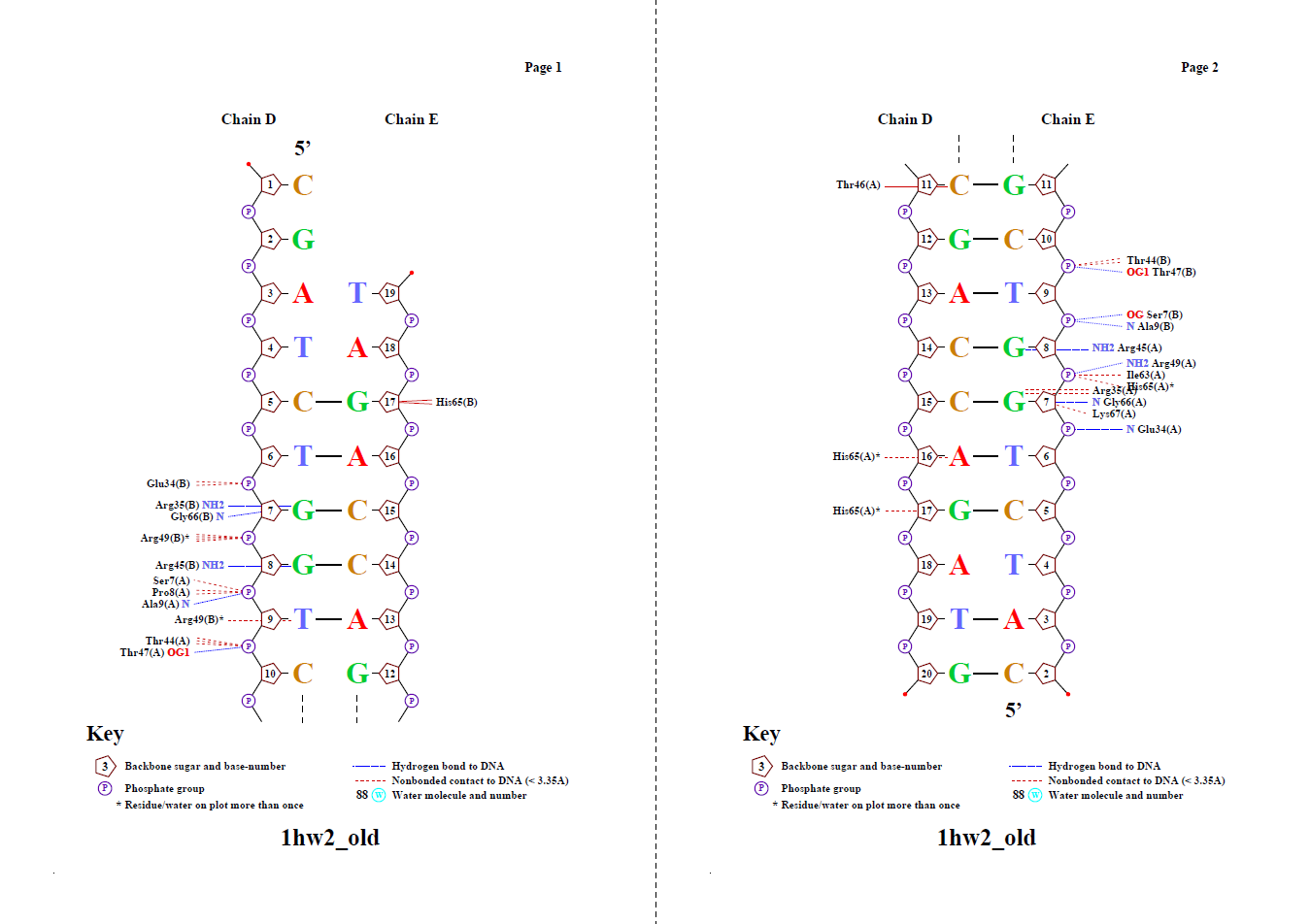 ♥
♥
A - наибольшее число контактов
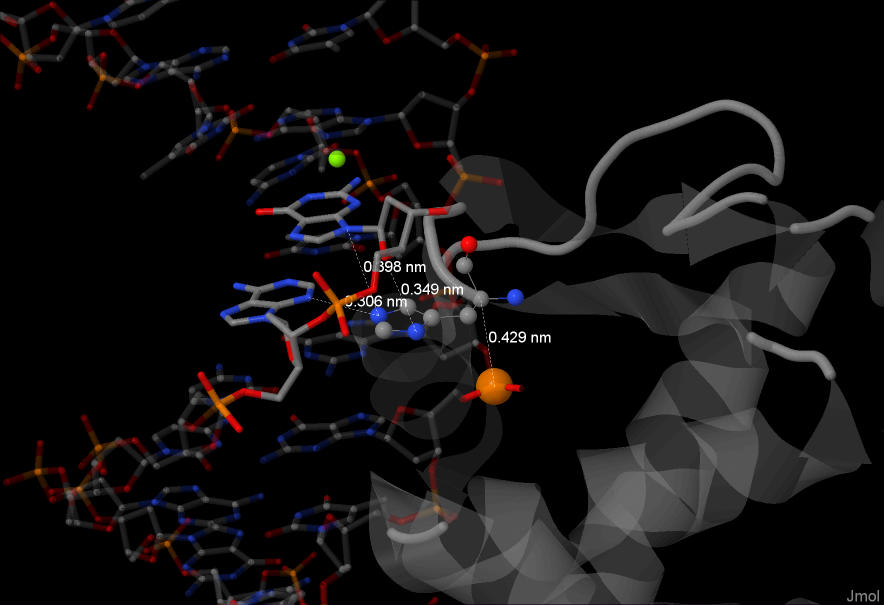
Наибольшее число контактов было найдено для гистидина №65 из цепочки А белка. Он взаимодействует с атомами 2х азотистых основанием а также с одним из
фосфоров остова.
На рисунке: белок серый, показана структура. Гистидин выделен цветом, представлен шариковой моделью.
ДНК в проволочной модели, полупрозрачная. Аденин (№16, цепочка D) и гуанин (№17, цепочка D), с которыми взаимодействует гистидин более плотной закраски.
Атом фосфора из остова цепочки Е выделен рыжим, большого радиуса. Посчитаны некоторые расстояния.
В - также важно для распознавания
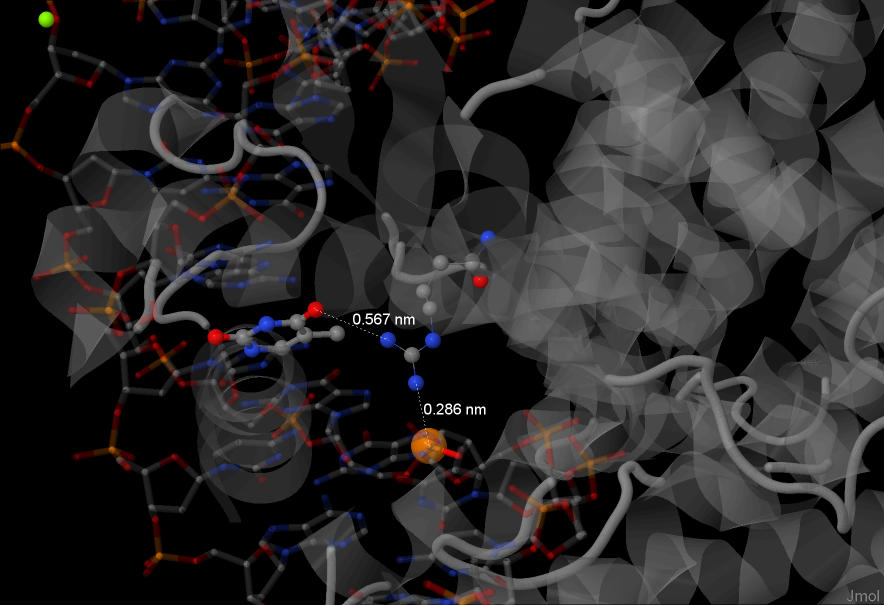
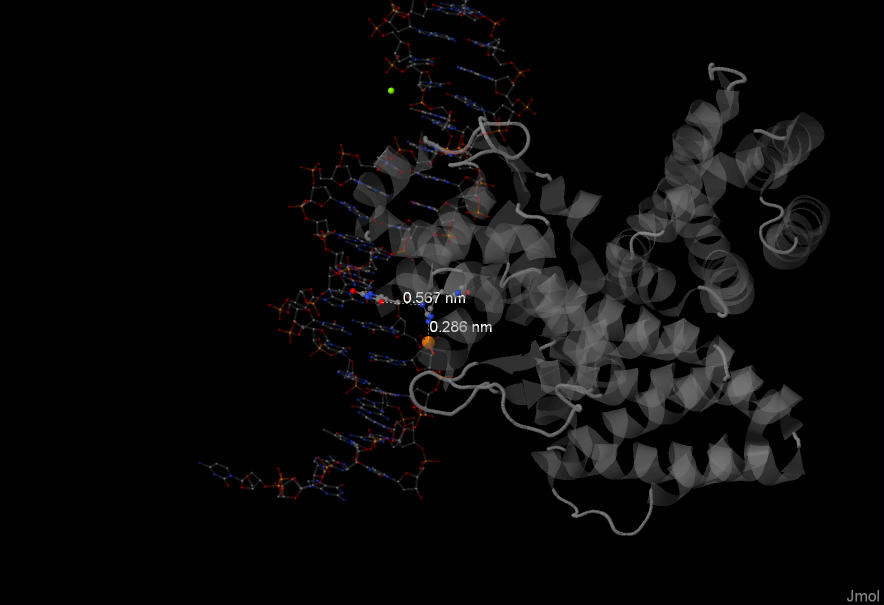
Также важным для распознавания ДНК, кажется, является аргинин №49 из цепочки В белка.
Он взаимодействует с тимином (№9, цепочка D) и с атомом фосфора. Расстояния достаточно близкие, что позволяет предположить специфичность взаимодействия.
Аргинин - основная аминокислота, его содержание высоко в гистонах - то есть аргинин в целом часто используется клеткой для контактов с ДНК.
Еще на отдалении в целом можно сказать, что данная область контакта +/- по центру всей области контактов. Возможно, он будет из-за этого реже разрушаться, или же там может находиться активный центр.
На рисунке: белок серый, показана структура. Аргинин выделен цветом, представлен шариковой моделью.
ДНК в проволочной модели, полупрозрачная. Тимин (№9, цепочка D) более плотной закраски.
Атом фосфора из остова цепочки D выделен рыжим, большого радиуса.