
В данном пункте были использованы белки из
Для сравнения было использовано несколько программ выравнивания: Tcoffee with Defaults, Probcons with Defaults, Muscle with Defaults и Mafft with Defaults, реализуемые
средствами програмы JalView, и программа COBALT:Multiply Alignment Tool-NCBI-NIH. Для сравнения были выбраны первая и последняя, из рассчёта, что сравнить было бы логично
аппарат, который чаще всего использовался в предыдущих работах(Tcoffee) и COBALT, условно не из цикла выравниваний JalView. К тому же первые четыре выравнивания достаточно
схожи, разумеется, в них есть отличия, и при детальном рассмотрении, наверное, наиболее отличается Mafft with Defaults,
но эти отличия не столь интересны и не столь иллюстративны.
Ссылки на исследуемые выравнивания так же приведены:
А чтобы посмотреть на весь проект, собственно, сравнения, целиком, кликнете на картинку(ссылка на скачивание):Начало выравниваний
Выравниваня покрашены Clustalx.Теперь, собственно, перейдём к отличиям выравниваний COBALT & TCoffee.
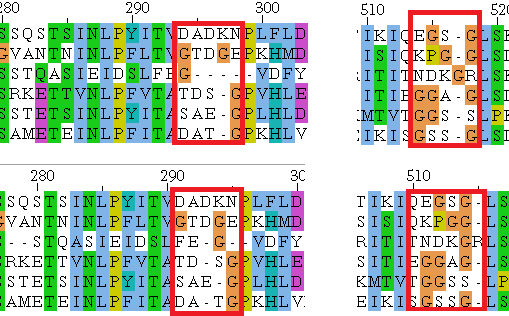
Фрагмент выравнивания
Наверное, это просто разные варианты прочтения, сложно сказать, как это регулируется.Для начала хотелось бы отметить, что Pfam не справился с поиском по UniProtID рабочего белка 4-alpha-glucanotransferase(AFH91336.1), из Corynebacterium pseudotuberculosis 31, и так как подобное не редкость, с этим белком в целом часто случались проблемы при поиске, как альтернативу не зазорно будет взять его гомолог, ранее найденный с помощью поиска BLAST - P65337.1|MALQ_MYCBO, который так же является 4-альфа-глюканотрансферазой Mycobacterium bovis AF2122/97. С ним проблем при поиске не возникло. <\p>
Данные по доменной организации белка представлены ниже: как можно пронаблюдать, MALQ_MYCBO имеет лишь один домен из Glycoside hydrolase family 77 Это достаточно большое семейство белков, как не сложно догадаться, их функция - гидролиз гликозидных связй между двумя или более карбогидратами или же между карбогидратом и какой-либо ещё молекулой. Для общего ознакомления так же отметим, что классификация насчитывает порядка ста семейств гликозид-гидролаз, эти данные основываются, очевидно, на различиях в сиквенсе.

Информация по доменной организации MALQ_MYCBO
При поиске по источникам Pfam нашлось 22 структуры, включающие достаточно приличное
количество различных архитектур, о которых хотелось бы рассказать ниже:
Можно сказать, что выпавший на мою долю белок имеет самую базовую комплектацию. Он содержит лишь один единственный домен и ничем особенно не примечателен.

Последовательностей такого типа насчитывается как-минимум 2094, как пример приведена самая первая архитектуру, хотя далее Pfam и предлагает при поиске очень схожие архитектуры, единственно что с наложением или отсутствием части выравнивания. Их было решено не рассматривать ниже. Зато к рассмотрению представлены архитектуры, которые содержат более-менее знакомые белки, чтобы разница в них была более наглядна.
Ещё две архитектуры похожего вида включают в себя домен альфа-амилазы. Когда мы говорим о "похожем виде" организаций, имеется виду визуальная составляющая, то есть наличие каких либо доменов в одинаковой последовательности. Разумеется, приведённые ниже архитектуры различаются положением доменов на цепи и предполагаемыми сайтами белкового связывания с альфа-амилазой, помеченными на рисунке: для первой структуры это 374 и 403 остатки, для второй: 356 и 385. К слову, на нижней структуре как раз можно видеть наложение частей последовательности, выравнены в структуре более тёмные части домена, хотя последовательность приведна, в данном случае, для него всего.
Ещё один вариант наличия альфа-амилазы в доменной архитектуре: это участок после интересующего нас домена гидролазы, претерпевший делецию концов своего участка.

Разумеется, альфа-амилазы тоже специфичны и могут быть представлены несколькими доменами:

Но в описании доменных архитектур так же хотелось бы отметить возможное наличие фософорилазы. Таких архитектур, как мы можем наблюдать, уже меньше, и домен фосфорилазы может быть представлен как отдельно от исследумого нами:


Небольщой оранжевый фрагмент перед гидролазой на первой структуре этого вида, видимо, сигнальный пептид.
И, напоследок, хотелось бы отметить возможное наличие глико-трансфераз: в данной архитектуры таковых присутствует целых две, последовательно после гидролазы.

Как можно наблюдать, функции рассмотренных выше доменов тесно связаны между собой, присутствие их в одной архитектуре не случайно:карбогидрат-фосфорилазы, например, используют тот же субстрат, что и рассматриваемые гилролазы. Далее приведена таблица с функциями доменов:
| Домен/Семья белков | Функция |
|---|---|
| Альфа-амилаза | Гидролизуют альфа-связи крупных полисахаридов |
| Карбогидрат-фосфорилаза | Cтимулируют образование глюкозы-1-фосфата из одного из гликогена, крахмала, глюканов или мальтодекстрин. |
| Гликозил-трансфераза | Катализируют перенос сахаридных фрагментов с "гликозил-донора" к нуклеофильной молекуле гликозил-акцептора |
Таблица 1. Функции доменов из различных архитектур, сопряжённых с исследуемым.
На главную страницуВернуться назад
©Solonovich Vera,2017