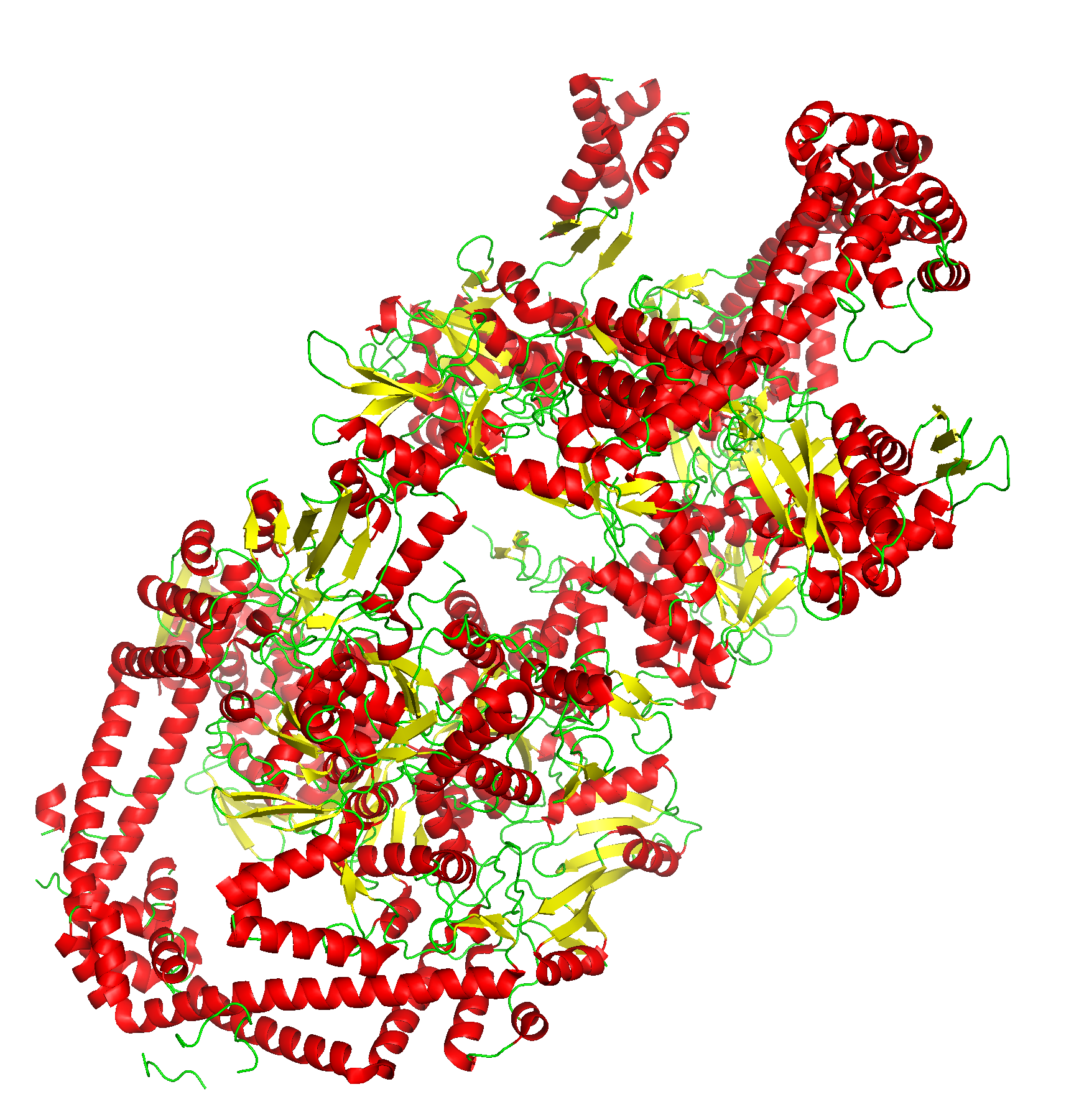


В качестве модеи была выбрана структура ДНК-гиразы из бактерии Escherichia coli.
Идентификатор PDB: 4TMA.
В поле CRYST1 соответствующего PDB-файла можно найти следующую запись-описание элементарной ячейки кристалла:
CRYST1 107.210 114.460 462.120 90.00 90.00 90.00 P 21 21 21 16Первые 3 числа обозначают длины сторон ячейки в Å (а, в и с соответственно, они же - направляющие векторы кристалла), следующие 3 - углы между ними, далее идет кристаллографическая группа P 21 21 21. Последнее число обозначает количество молекул в ячейке.
|
|
|
| a | б | в |
Рисунок 1. ДНК-гираза. а. Ассиметрическая единица структуры. б, в. Комплекс ДНК-гиразы А2В2, А-субъединицы выделены зеленым, В-субъединицы - красным
Для заполенения элементарной ячейки структурами в PyMol была использована команда symexp:
symexp crystall, 4tma, 4tma, 30То есть,мы строим повторяющиеся объекты 4tma, удаленные не более чем на 30 Å от объекта 4tma. В результате были получены изображения укладки молекул в кристалле (Рис.2).
 |
 |
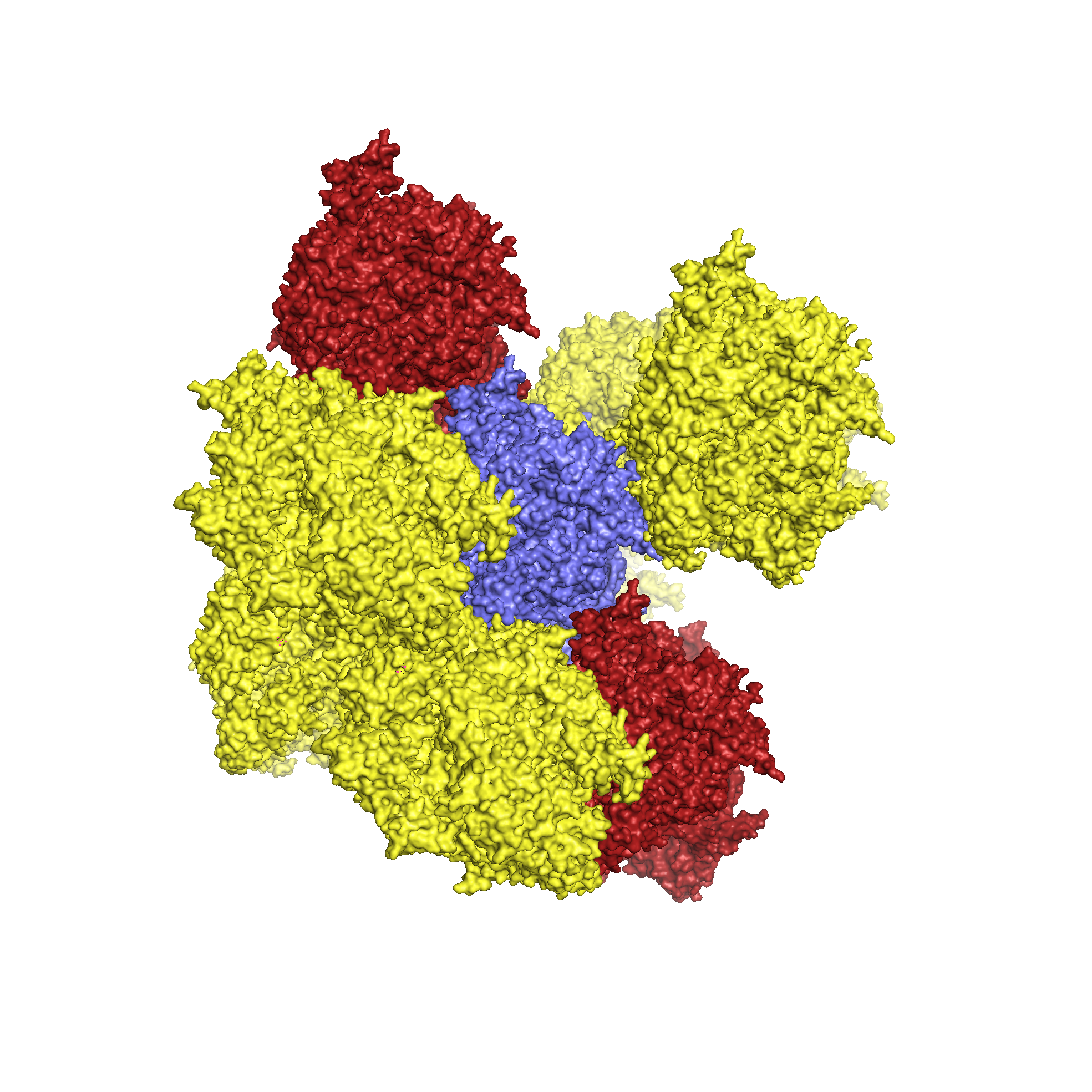 |
Рисунок 2. Укладка молекул ДНК-гиразы в кристалле, вид с разных ракурсов. Для наглядности, молекулы раскрашены в разные цвета.
Укладка молекул в кристалле не очевидна,- слои молекул как бы повернуты относительно друг друга. Если увеличить область построения кристалла до 50 Å вокруг исходной молекулы, то картина станет немного яснее, по крайней мере более понятно представлены слои молекул (Рис.3а). Если же добавить параллелепипед элементарной ячейки командой show cell, то можно себе представить, как в него вписаны молекулы (Рис.3б). Честного заполнения ячейки структурами провести не удалось, так-как обработка большого количества субъединиц требует большей мощности, и компьютер плохо справляется с этой задачей. |
 |
| a | б |
Рисунок 3. Укладка молекул ДНК-гиразы в кристалле. а. Слои молекул в кристалле. б. Положение элементарной ячейки в кристалле, вид вдоль длинейшего вектора с
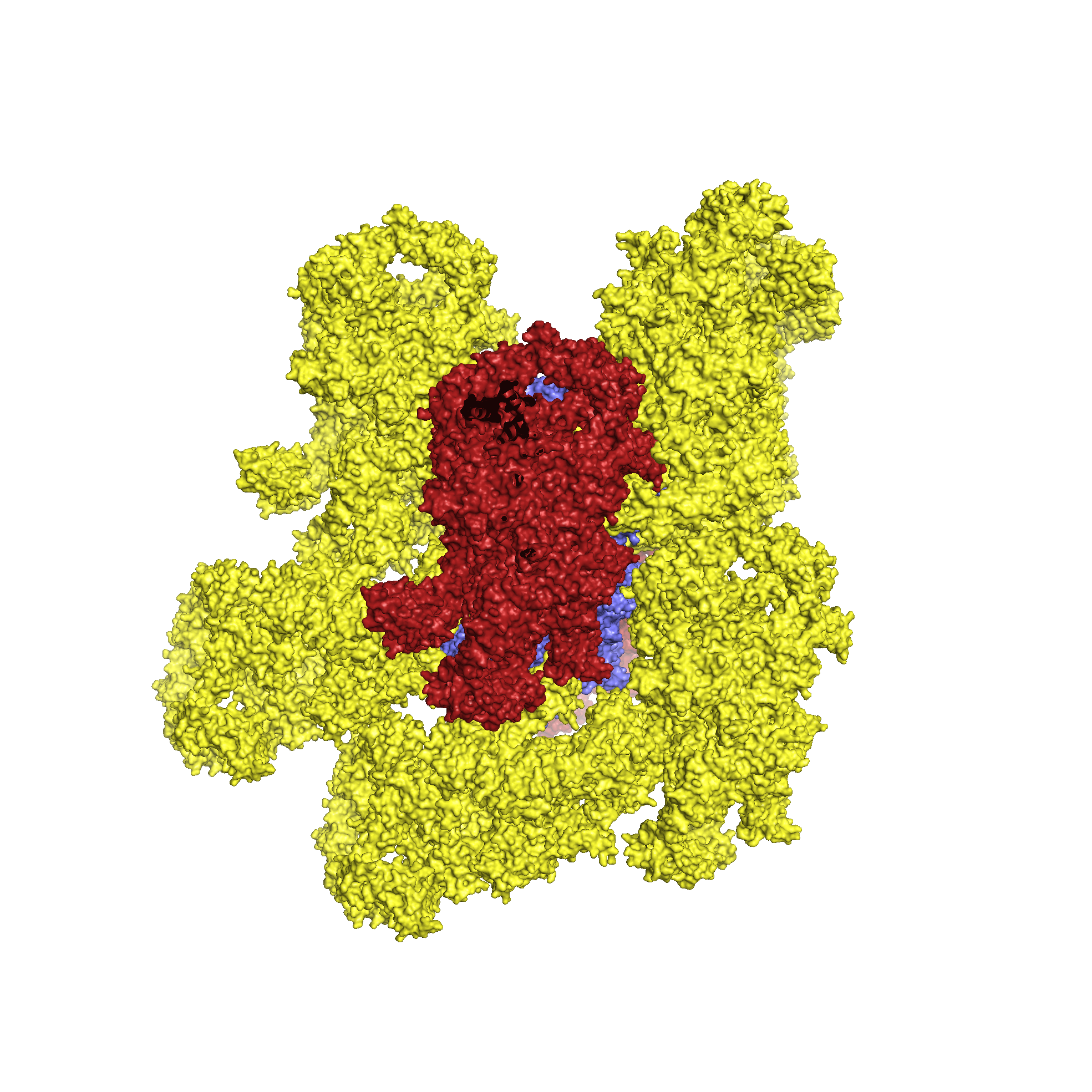
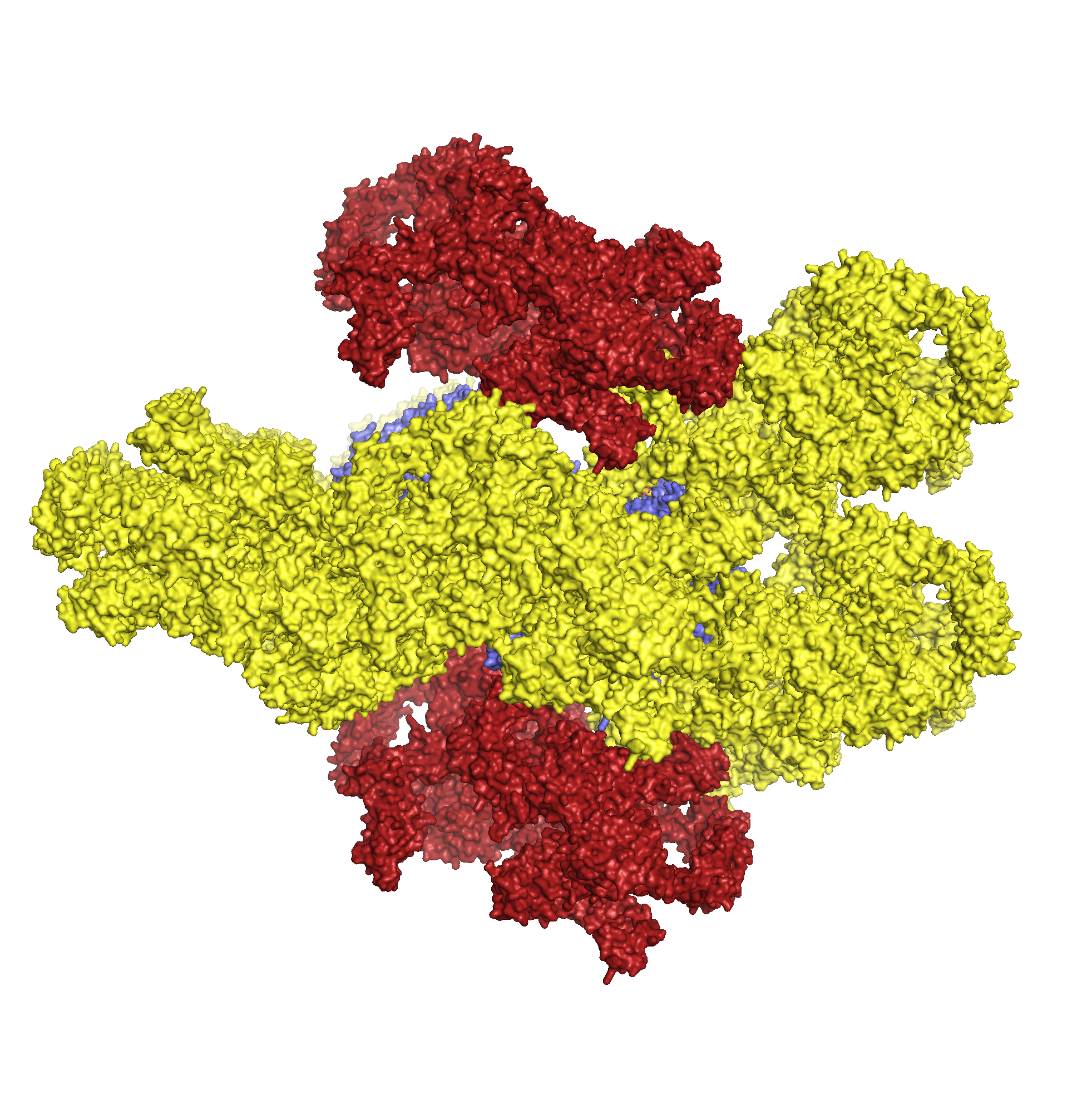
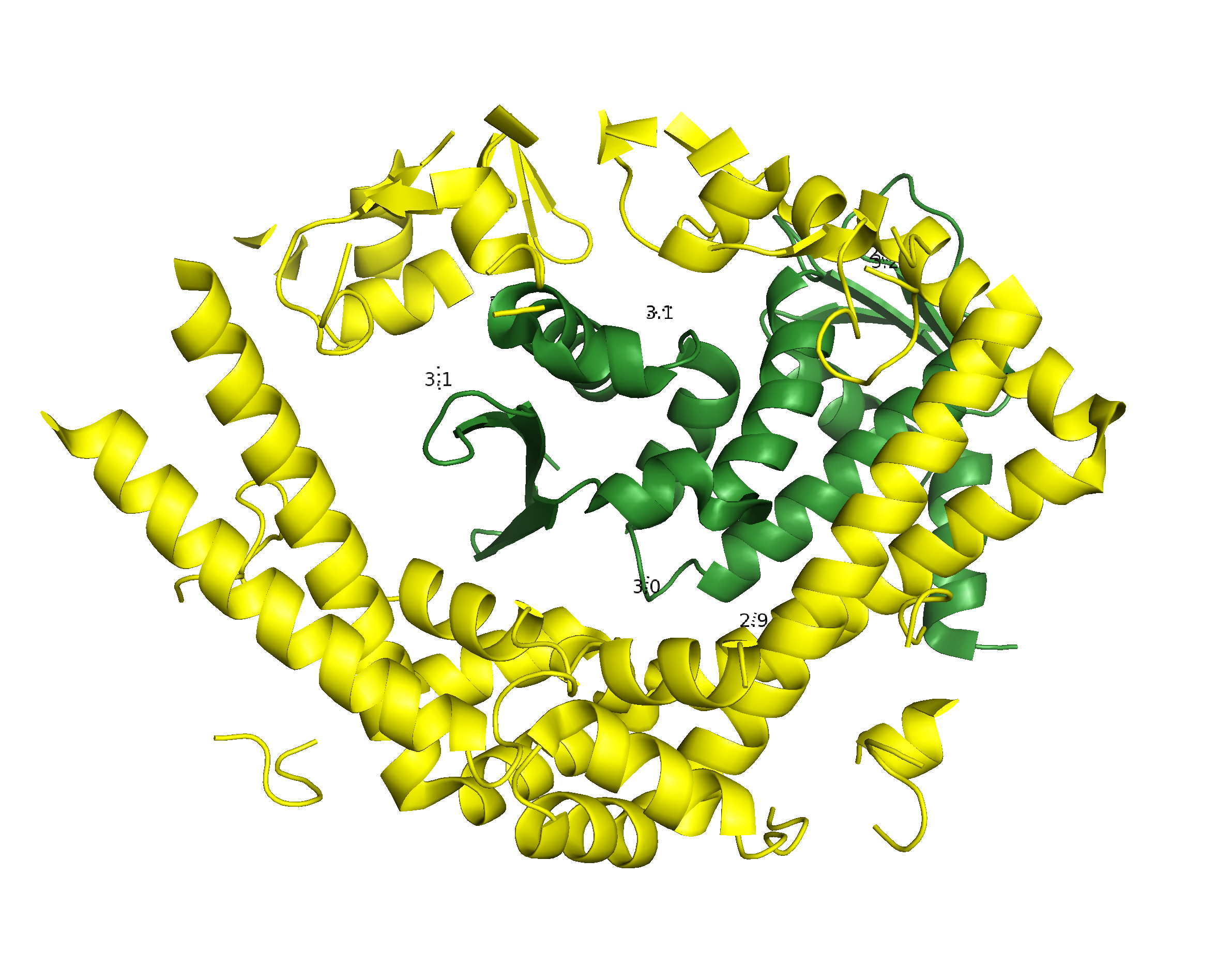
Для исследования контактов между молекулами в кристалле была создана струткура из трех пар ДНК-гираз (Рис.4, центр), посредством команды symexp с параметром "близости" молекул - 2 Å. В полученной структуре можно выделить две принципиально разные зоны контактов (Рис.4, контакт типа 1 и типа 2). При этом, контакт типа 2 присутствует в двух экземплярах.
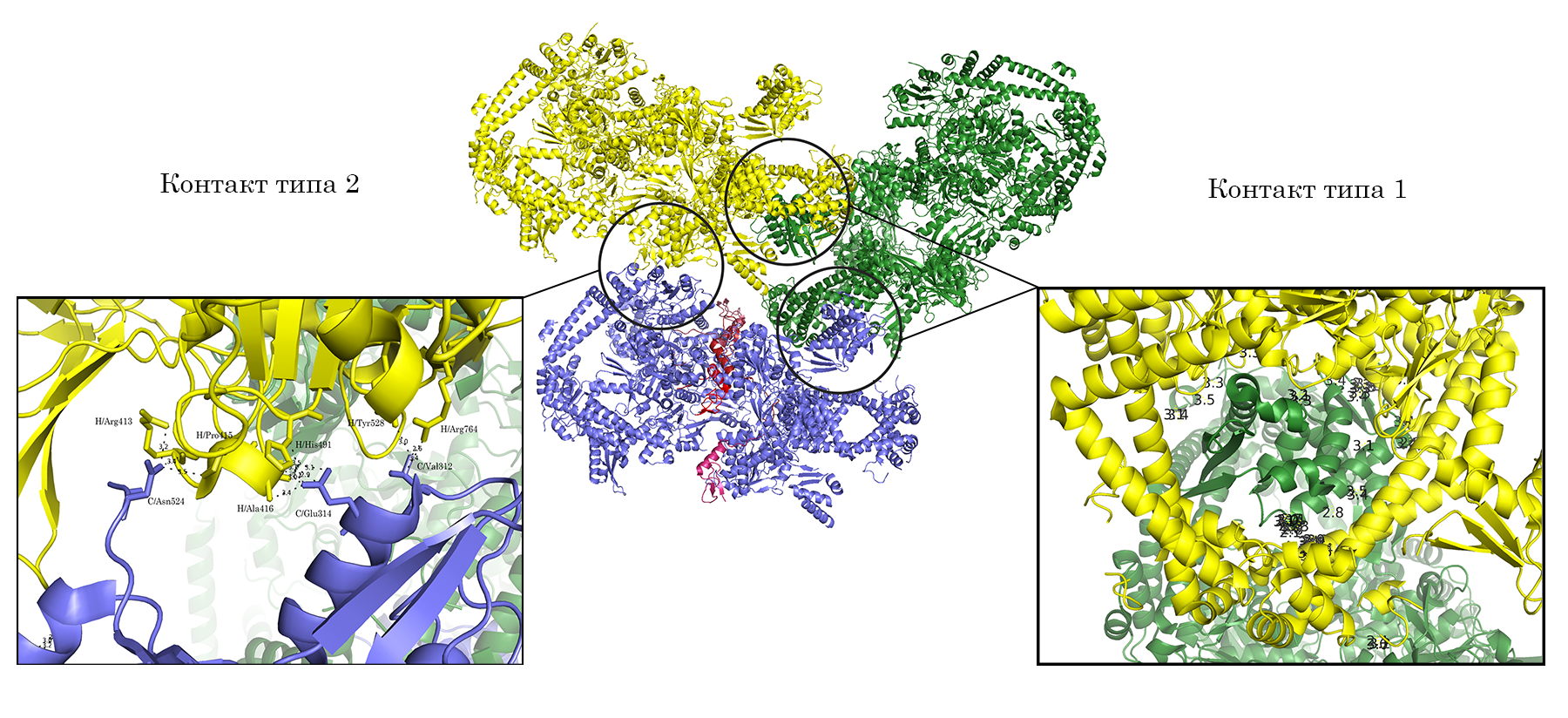
Рисунок 4. Ассиметрические единицы, расстояние между которыми не больше 2 Å и общий вид двух областей контакта между соседними единицами.
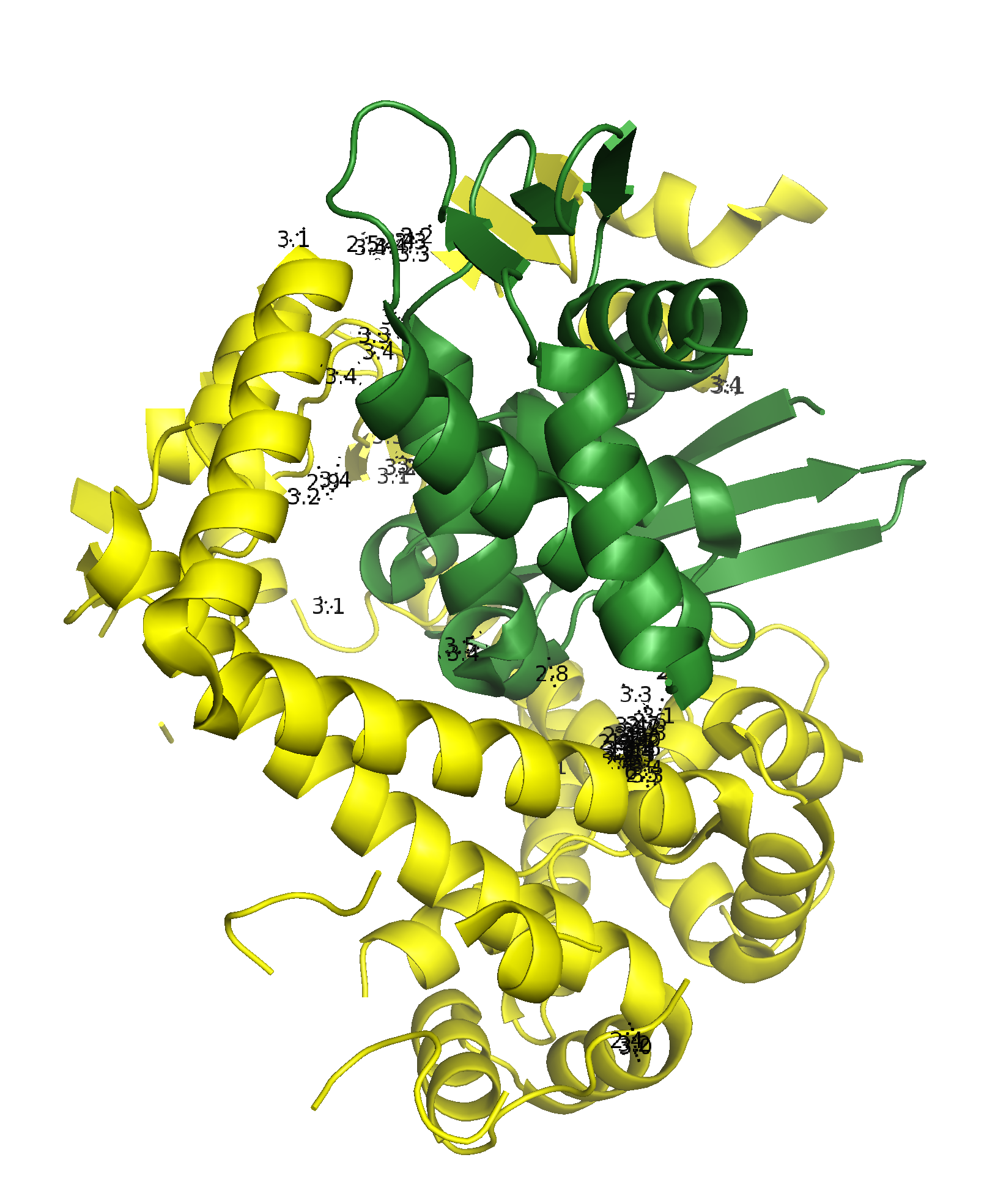
Подвижный домен субъединицы В практически заходит в кольцо, образованное А-субъединицами соседней молекулы, образуя контакт типа 1 (Рис. 5).
 |
 |
| a | б |
Рисунок 5. Общий вид контакта типа 1. а.Вид сквозь кольцо А-субъединиц на домен В-субъединицы соседней молекулы. б. Вид контакта сбоку; между молекулами отмечены расстояния не большие, чем 3.5 Å
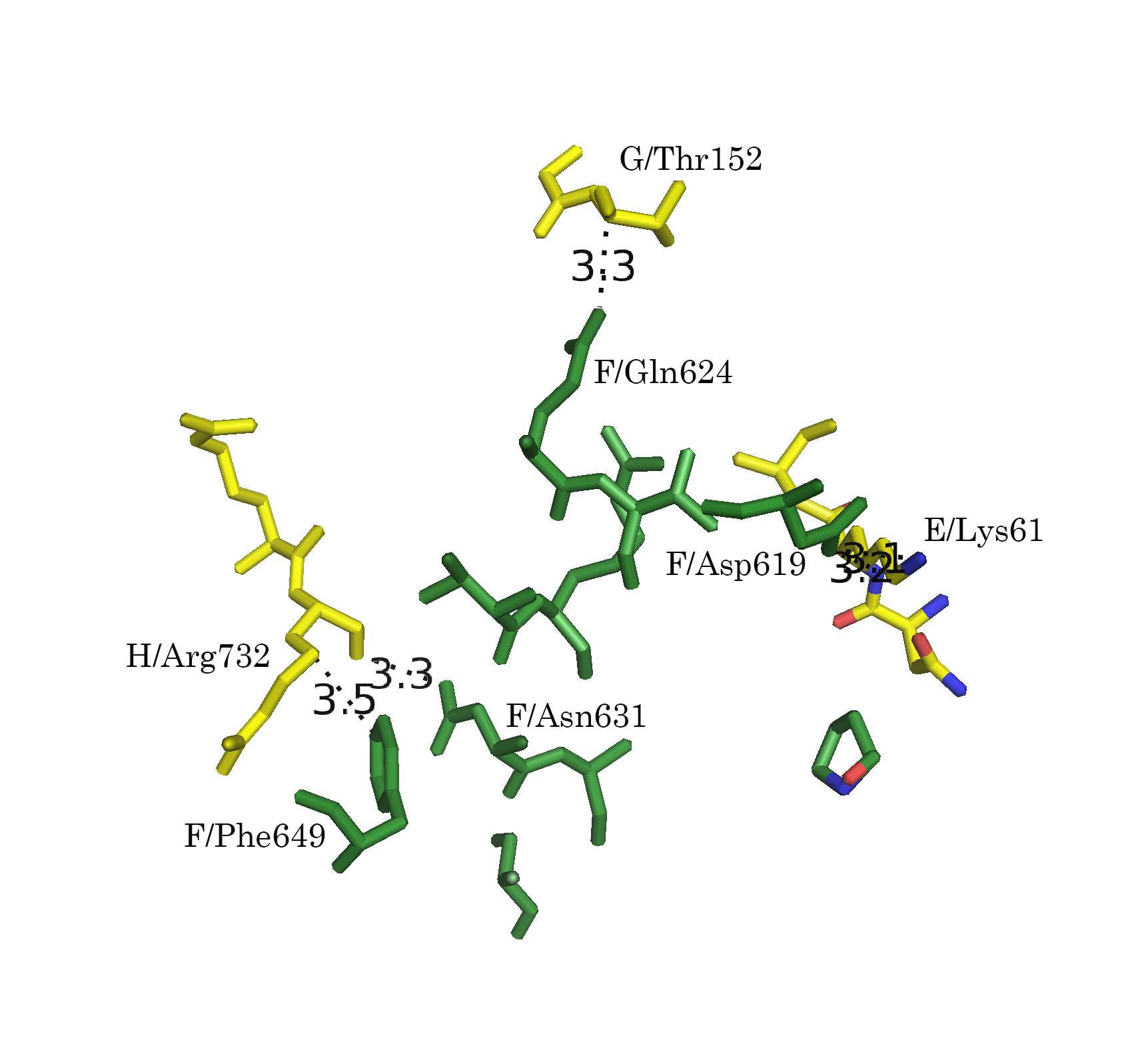
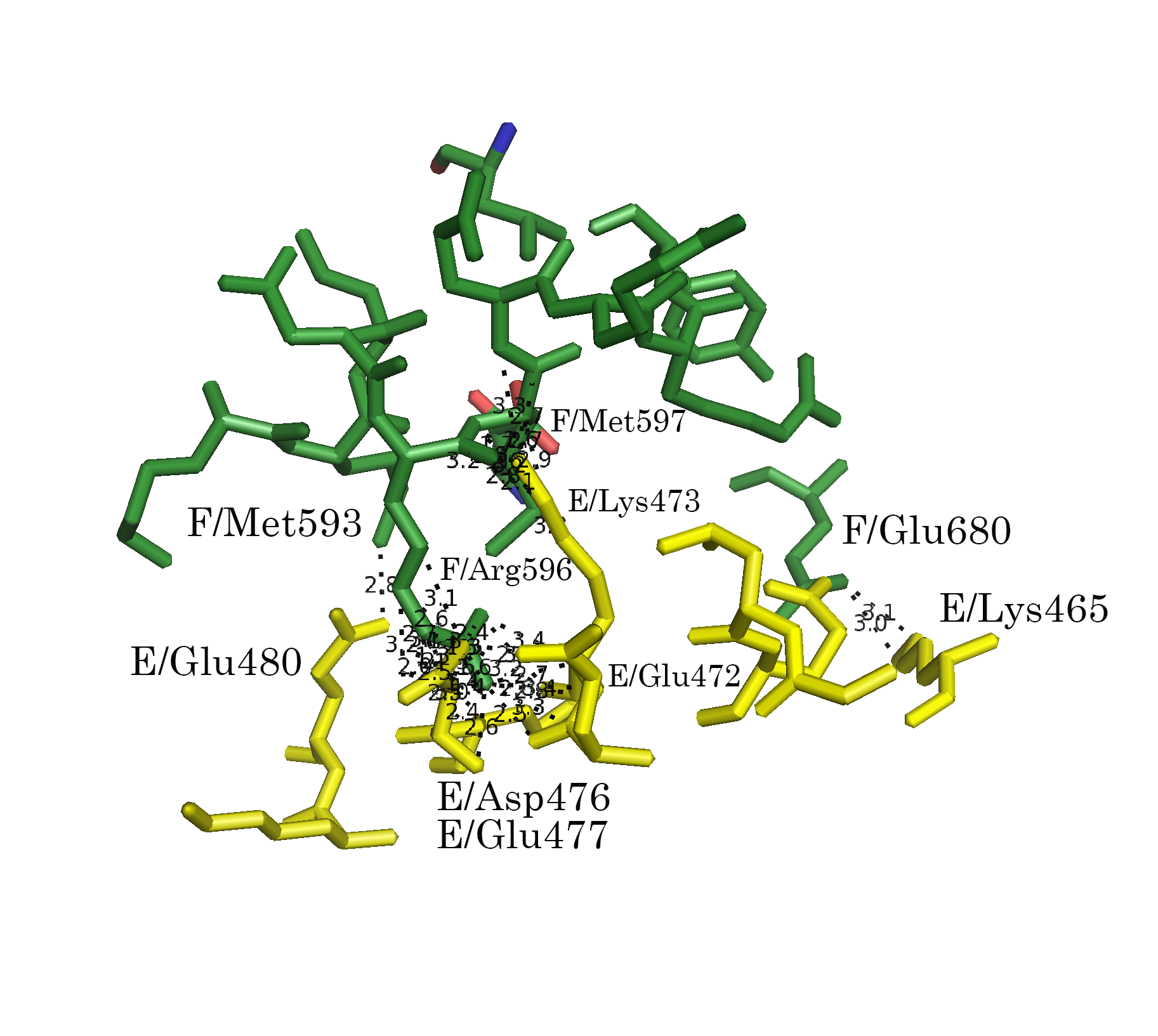
Командой distance, byres были показаны расстояния между всеми аминокислотными остатками разных ассимметрических единиц, не превышающие 3.5 Å:
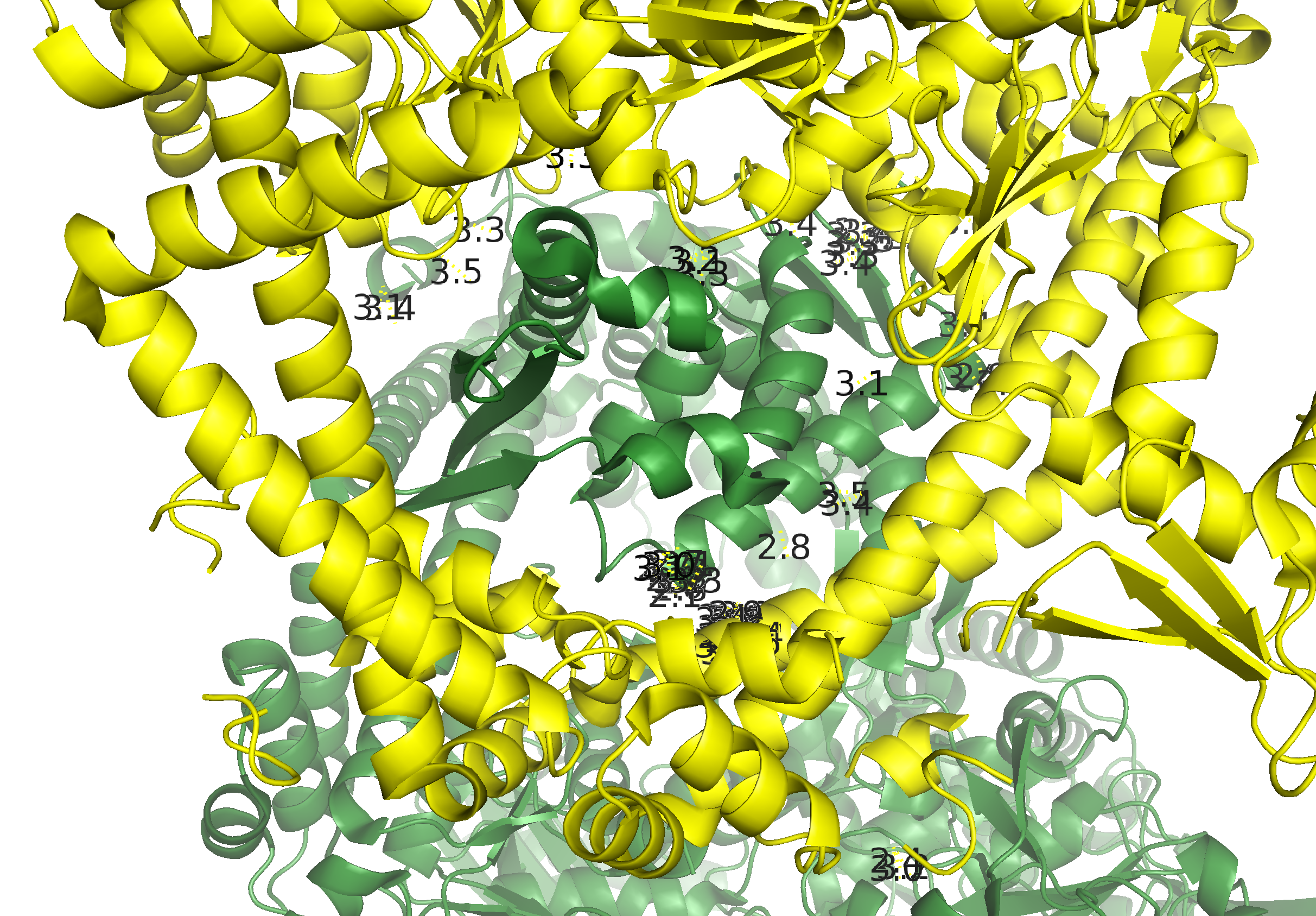
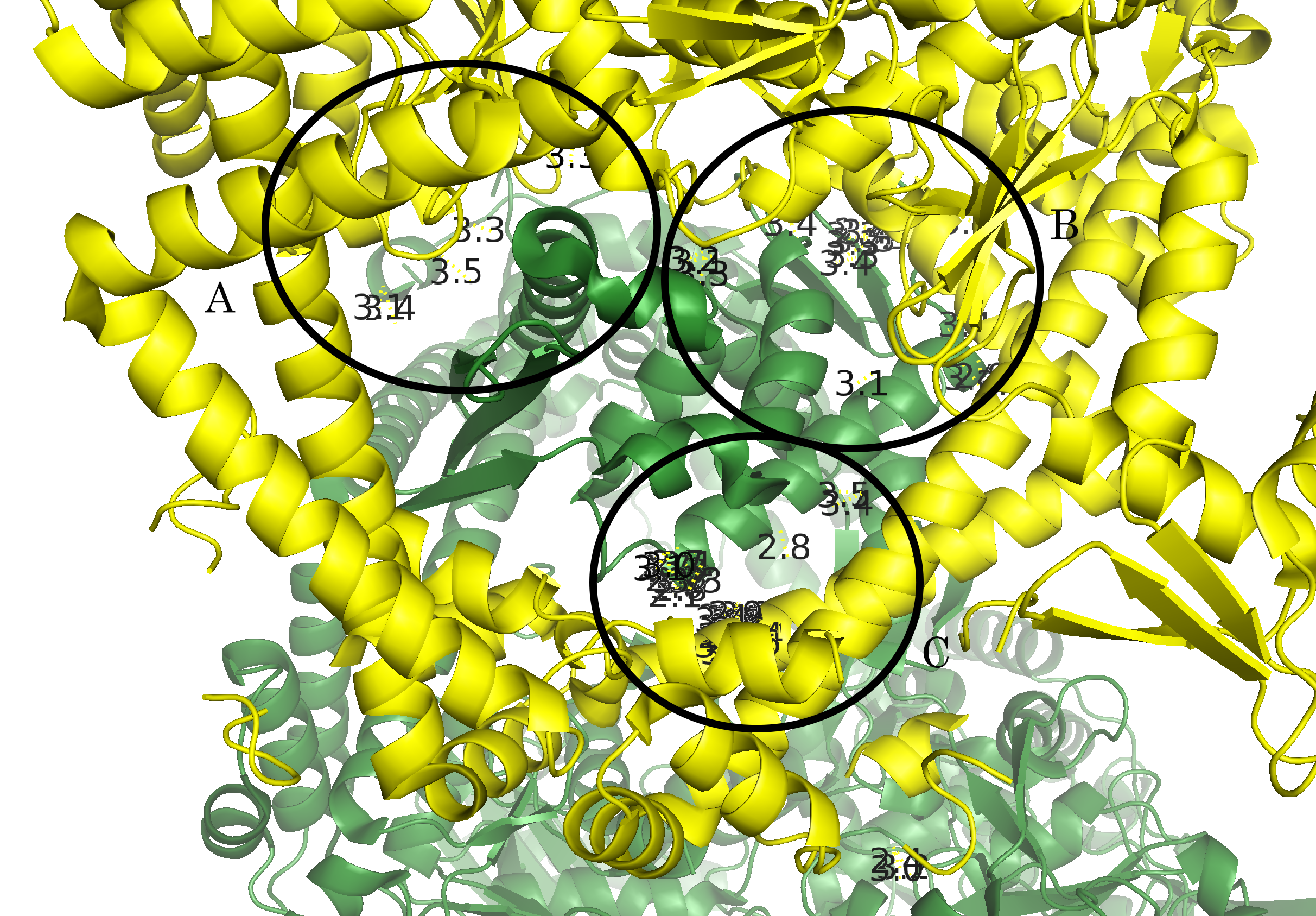
distance bonds_1, 4tma, sym0300-100, 3.5Из рисунка 5а видно, что контакты расположены практически по всему периметру кольца А-субъединиц. Для более детального описания контактов, вся область условно была разбита на 3 кластера контактов (Рис. 6а): А, В, С (Рис. 6б-г).
 |
 |
 |
 |
| a | б | в | г |
Рисунок 6. Контакт типа 1. а.Общий вид контакта с выделенными кластерами контактов. б. Область А. в. Область В. г. Область С; расстояния указаны в Å
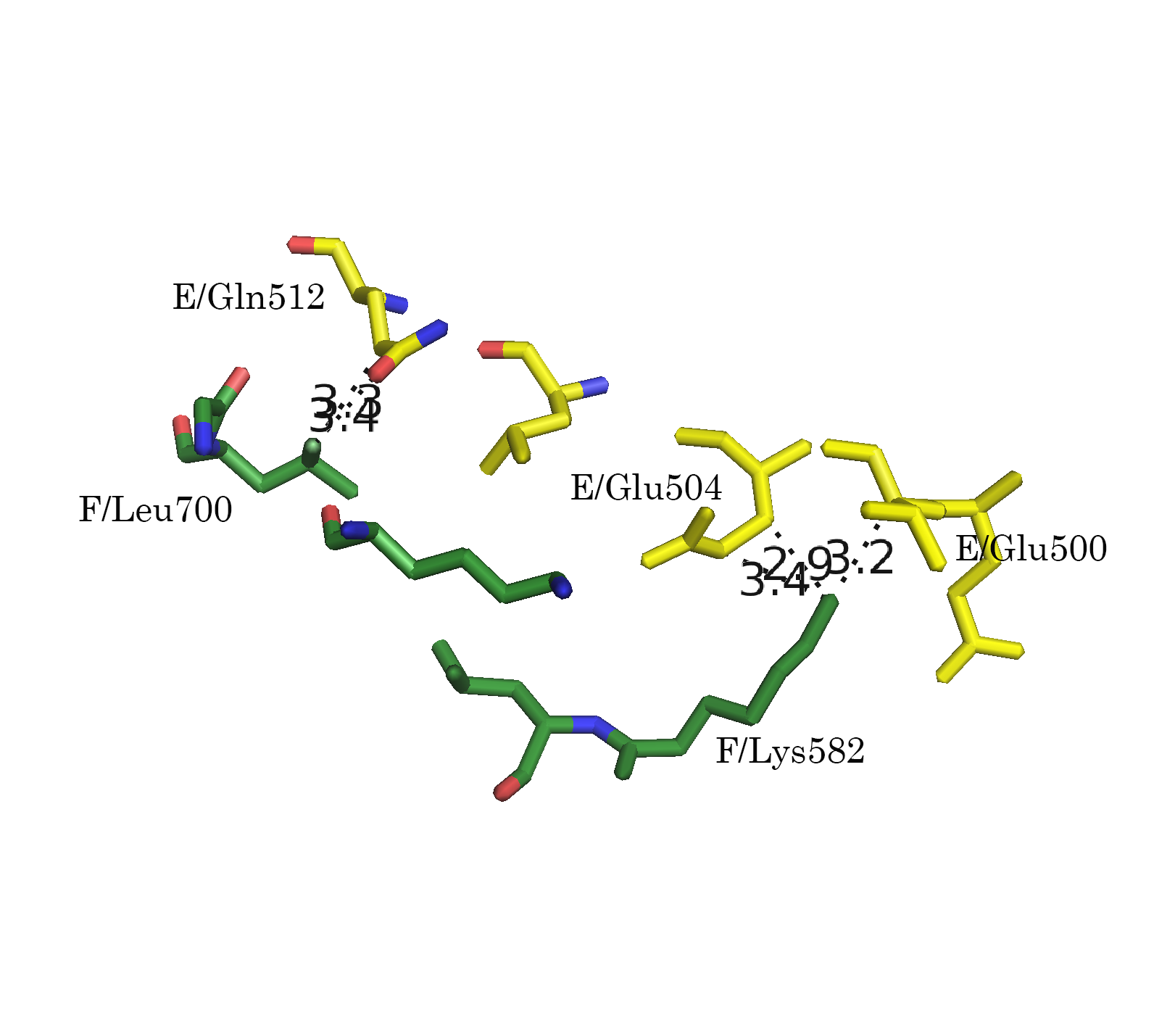
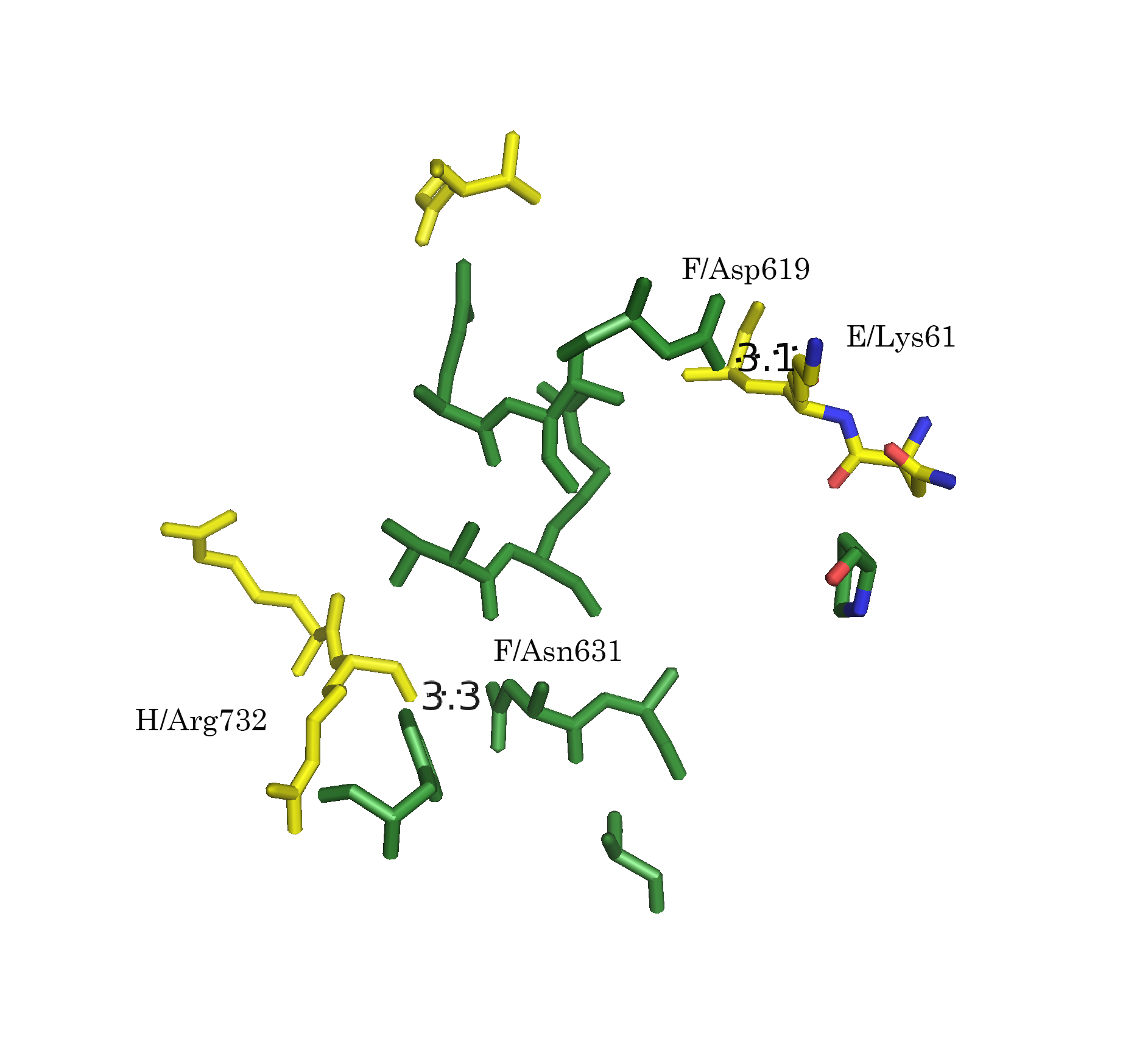
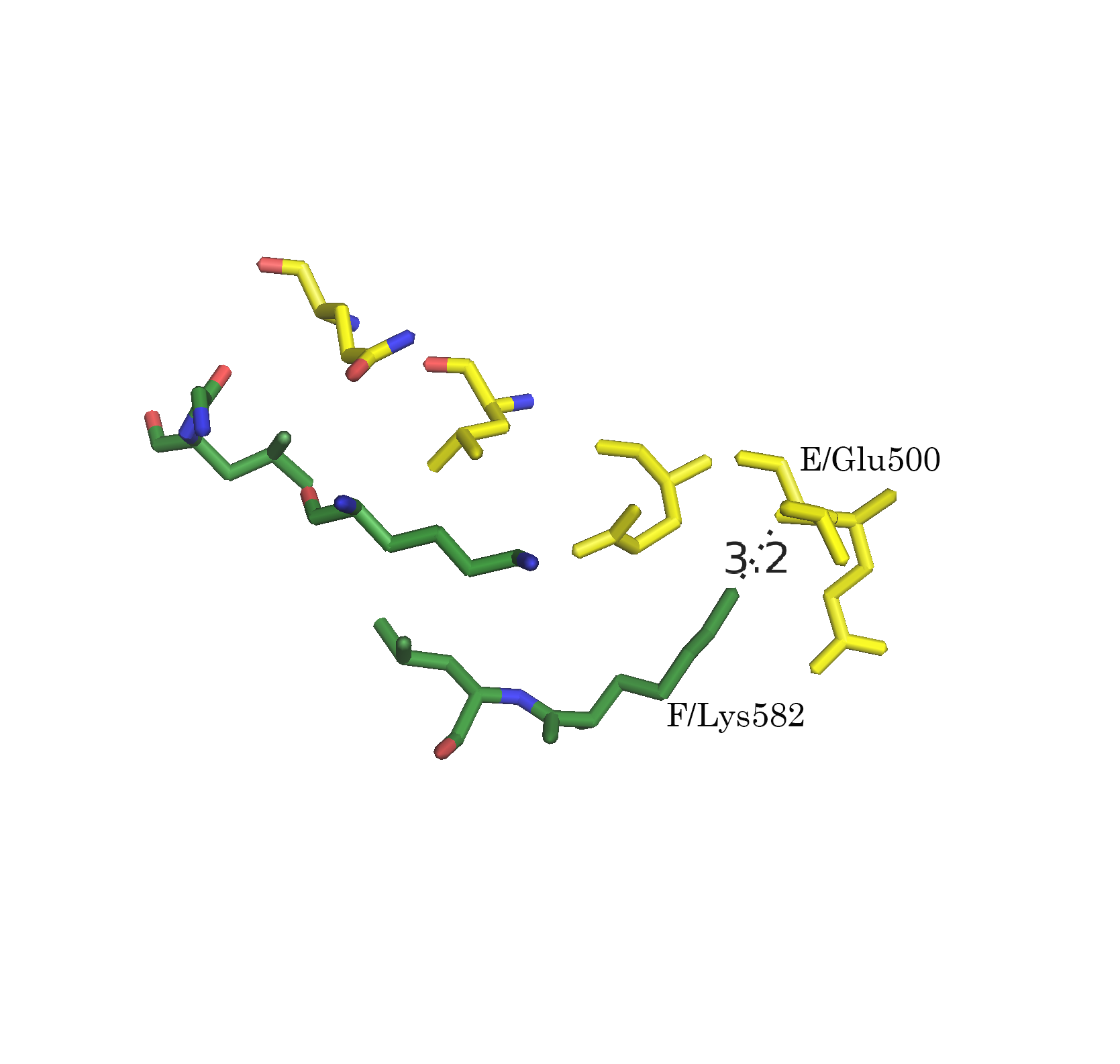
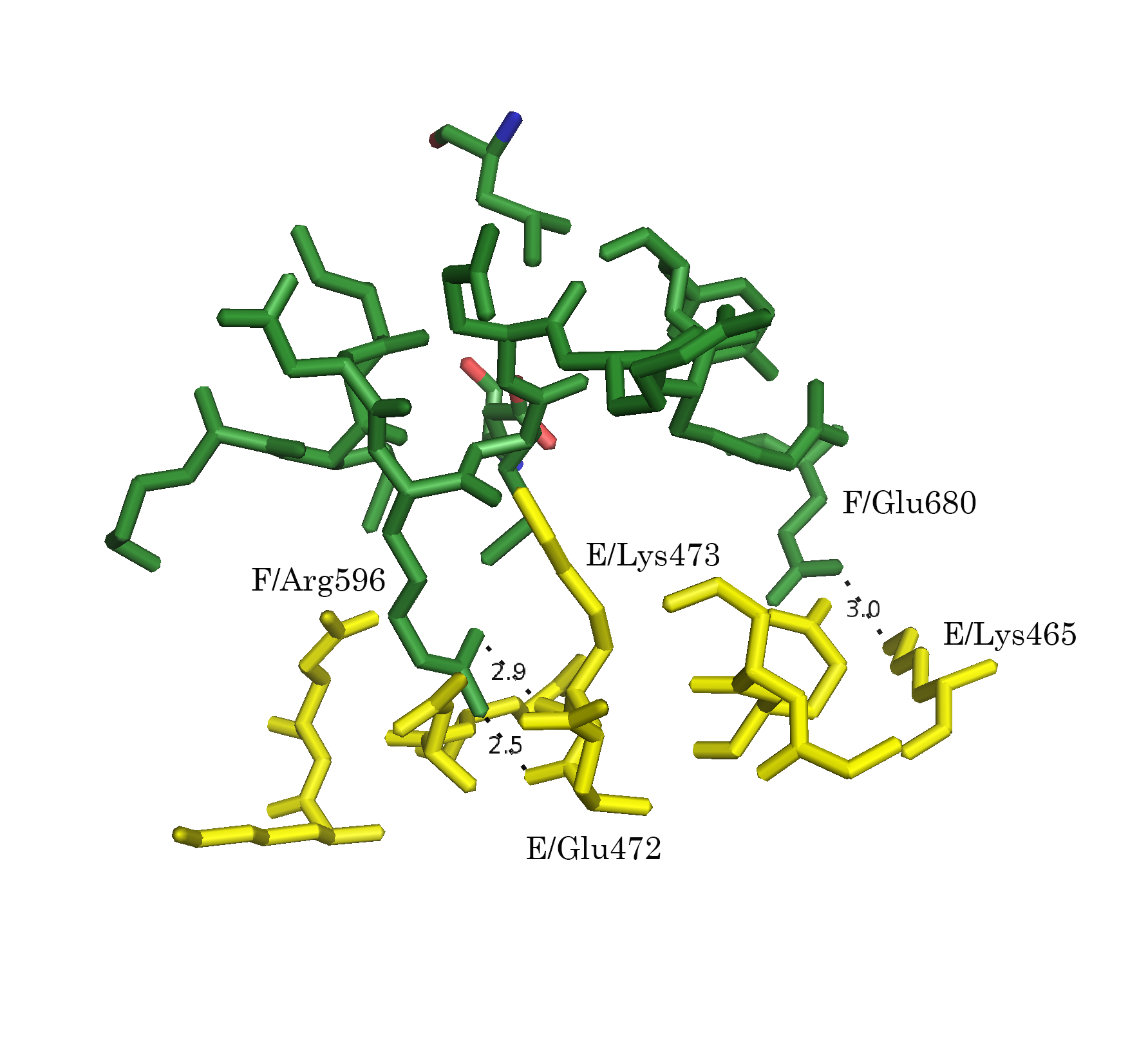
Следует отметить, что функция distance, byres без дополнительных параметров размечает любые расстояния. Для того, чтобы получить данные только для полярных контактов (подозрения на водородные связи, ионные взаимодействия), следует указывать флаг 2:
distance name, sel1, sel2, 3.5, 2После применения указанной модификации, количество размеченых контактов упало (Рис. 7а-г).
 |
 |
 |
 |
| a | б | в | г |
Рисунок 7. Полярные взаимодействия в области контакта типа 1. а.Общий вид контакта. б. Область А. в. Область В. г. Область С; расстояния указаны в Å
Обращает на себя внимание взаимодействие 596 Arg B-субъединицы с А-субъединицей в области С (Рис. 6г и 7г). Этот положительно заряженный остаток встроен в отрицательно заряженную "корзину", состоящую из 476 Asp, 477 Glu и 472 Glu.
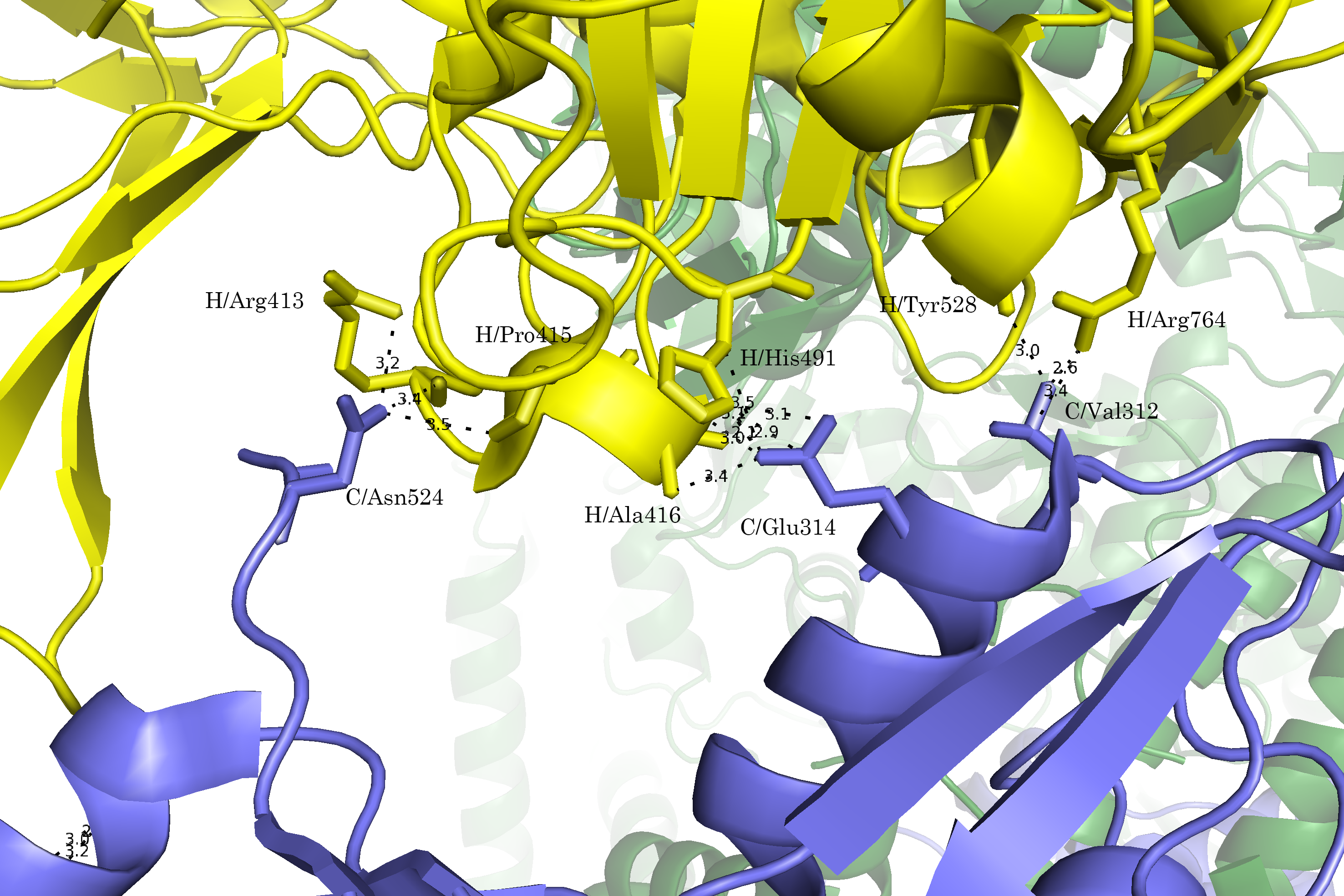
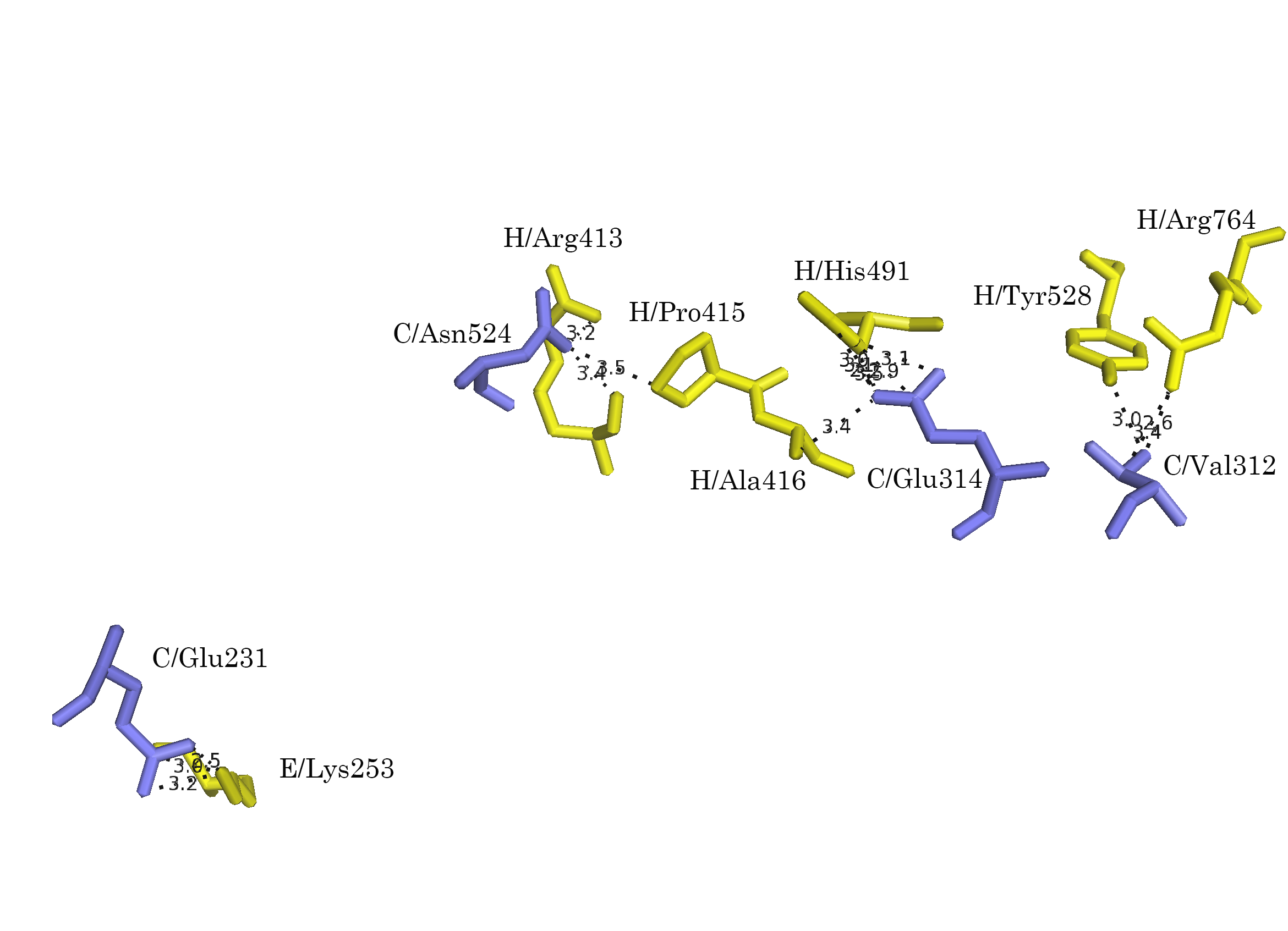
Контакт 2 образован боковыми С-концевыми доменами А-субъединиц разных молекул. Для контакта были проведены все те же действия, что и для первого случая. Были выявлены сближенные аминокислотные остатки и показаны потенциальные полярные взаимодействия (Рис. 8а-г).
 |
 |
 |
| a | б | в |
Рисунок 8. Область контакта типа 2. а.Общий вид контакта. б. Сближенные аминокислотные остатки. в. Полярные взаимодействия; расстояния указаны в Å
Для ДНК-гиразы, насколько это известно из литературы, олигомеризация не характерна. С другой стороны, фермент в процессе своей работы претерпевает очень сильные конформационный перестройки, такие как сближение доменов В-субъединиц, размыкание кольца, образованного А-субъединицами и тому подобные. Возможно, контакты в кристалле показывают взаимодействия между частями фермента в его альтернативных рабочих конформациях.
Для анализа расположения белковых цепей была выбрана структура ДНК-белкового комплекса (PDB ID: 3HDD), представляющего собой транскрипционный фактор Engrailed, связанный с ДНК (Рис. 9). Этот белок содержит гомеодомен и принимает участие в развитии дрозофилы. Гомеодомен – это тип белковой укладки "спираль-поворот-спираль" из 60 аминокислотных остатков. Он состит из трех соединенных между собой короткими петлями альфа-спиралей, одна из которых связывает консервативный сайт TAAT.
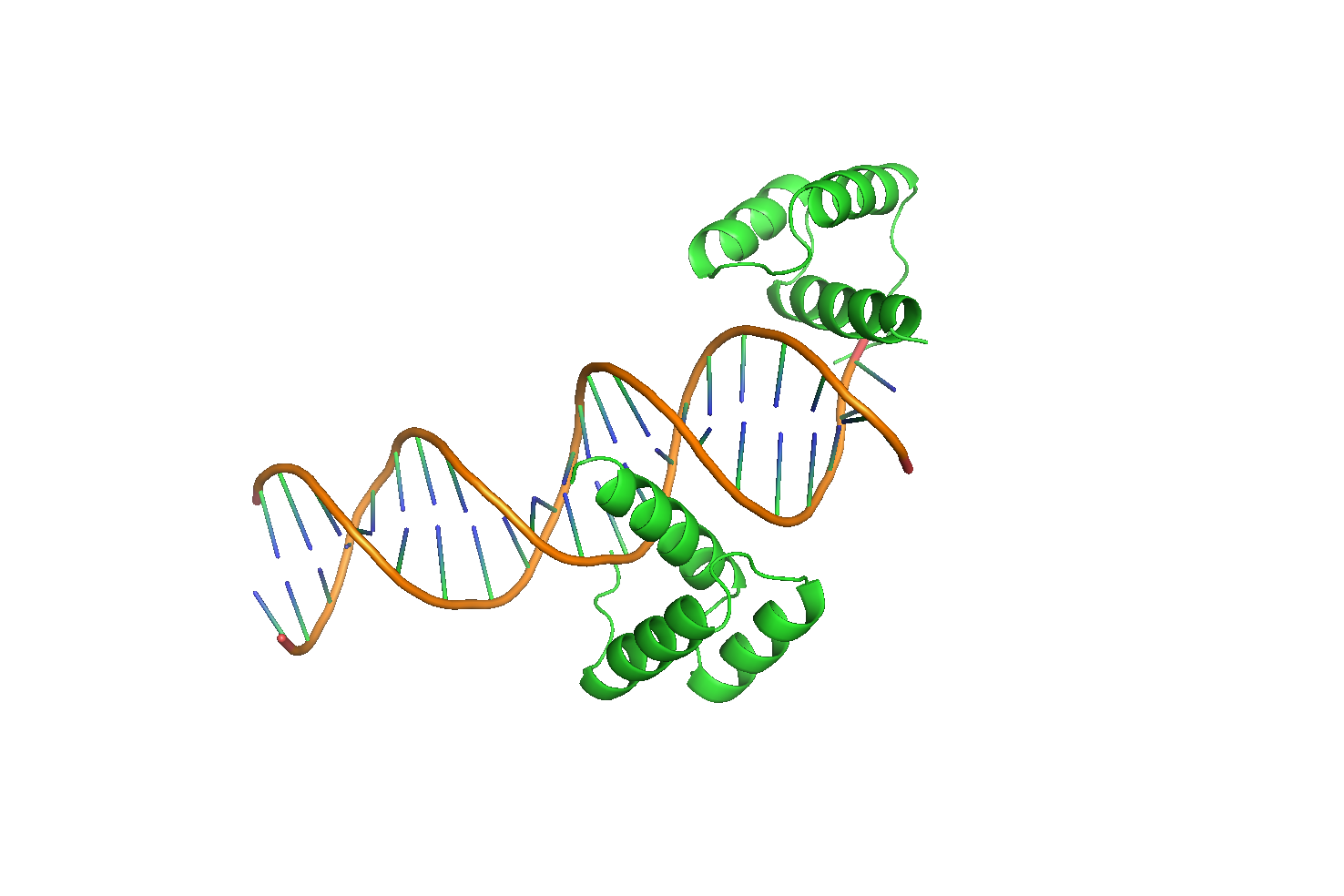
Рисунок 9 Структура 3HDD
Если применить команду symexp, то можно увидеть расположение других молекул в кристалле (Рис. 10):
symexp add, 3hdd, 3hdd, 5
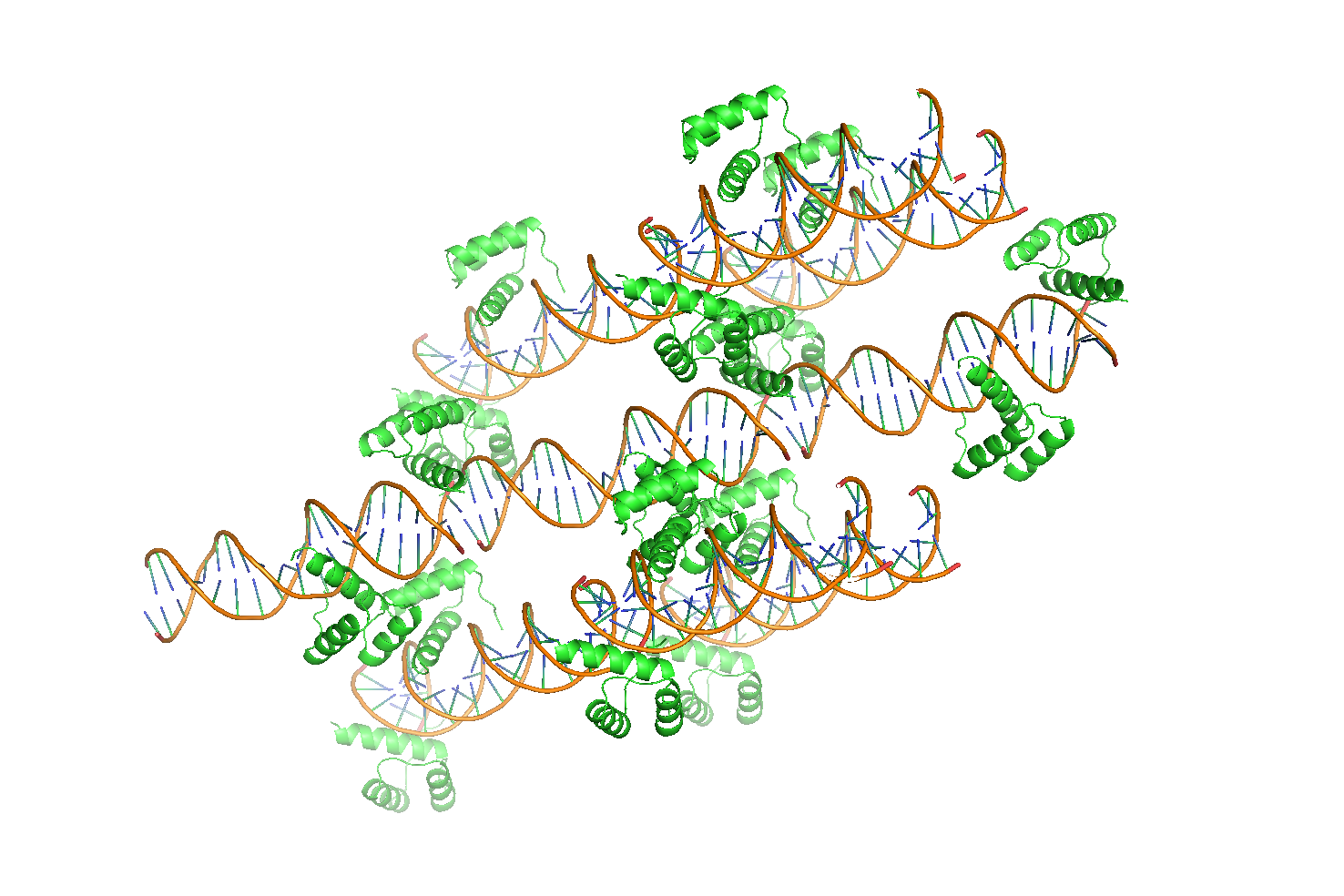
Рисунок 10 Структура кристалла 3HDD
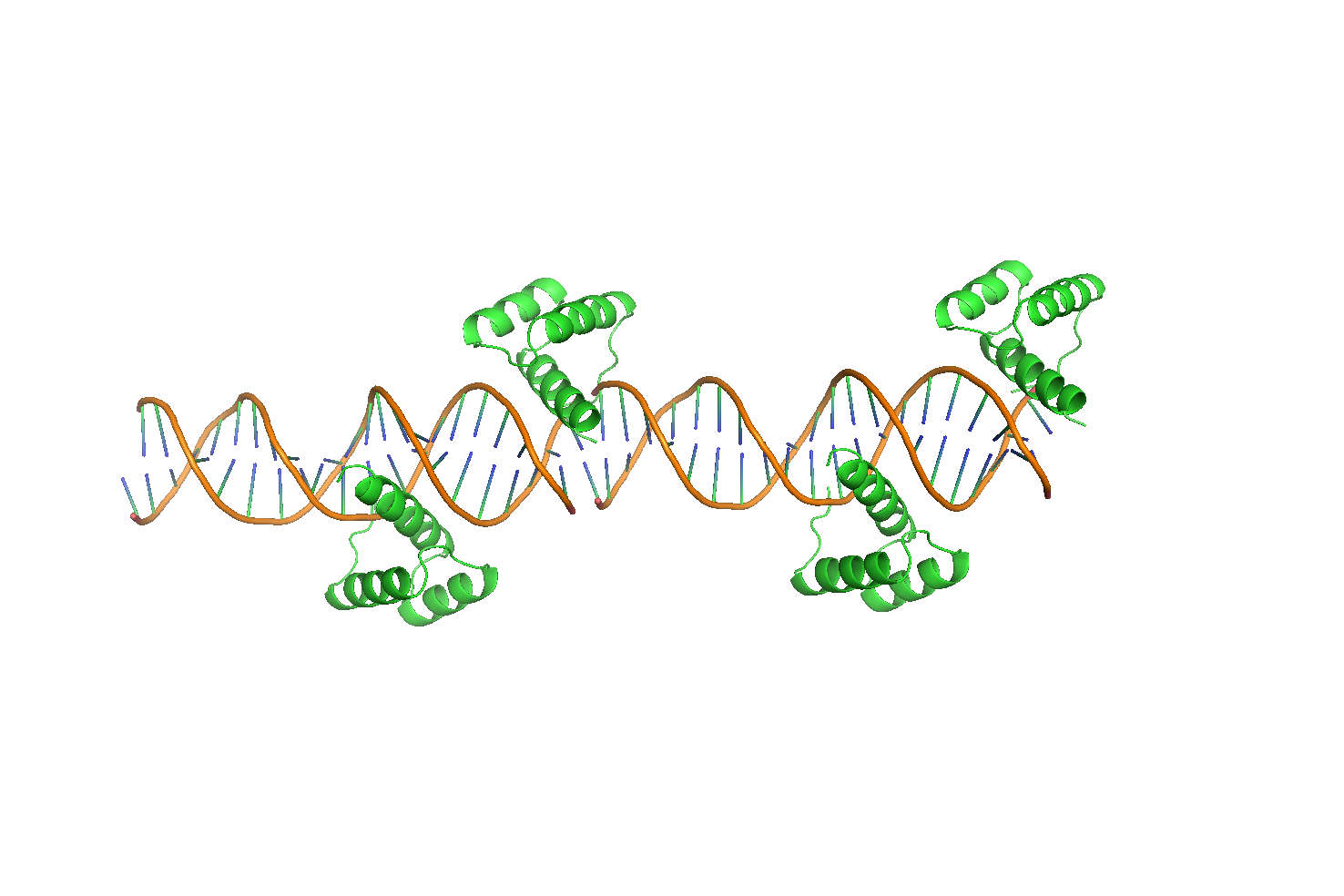
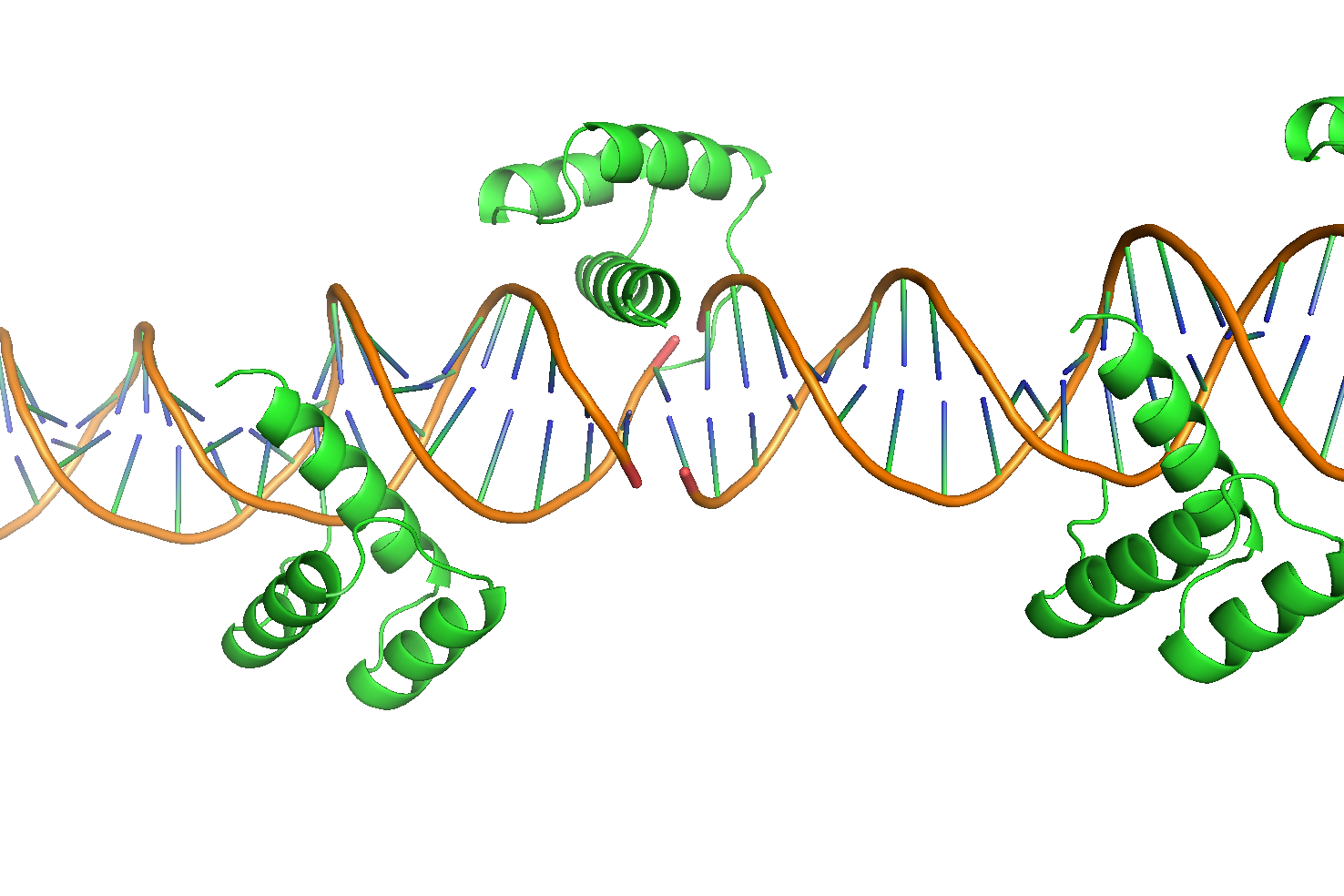
Интересно, что фрагменты ДНК выстраиваются линейно друг за другом. Для выяснения причины такого поведения, было получено изображение двух последовательно соединенных участков (Рис. 11a).
 |
 |
| a | б |
Рисунок 11 Две, последовательно расположенные, структуры 3HDD. а. Общий вид "дуплекса"б. Зона контакта двух фрагментов ДНК.
Можно видеть, что в гомеодомен, расположенный в торце первого фрагмента, взаимодействует также и со второй молекулой ДНК (Рис. 11б). Более того, фрагменты ДНК несут липкие концы из одного нуклеотида, которые очевидно взаимодействуют друг с другом и таким образом наращивают дуплекс ДНК фрагмент за фрагментом (Рис. 12).
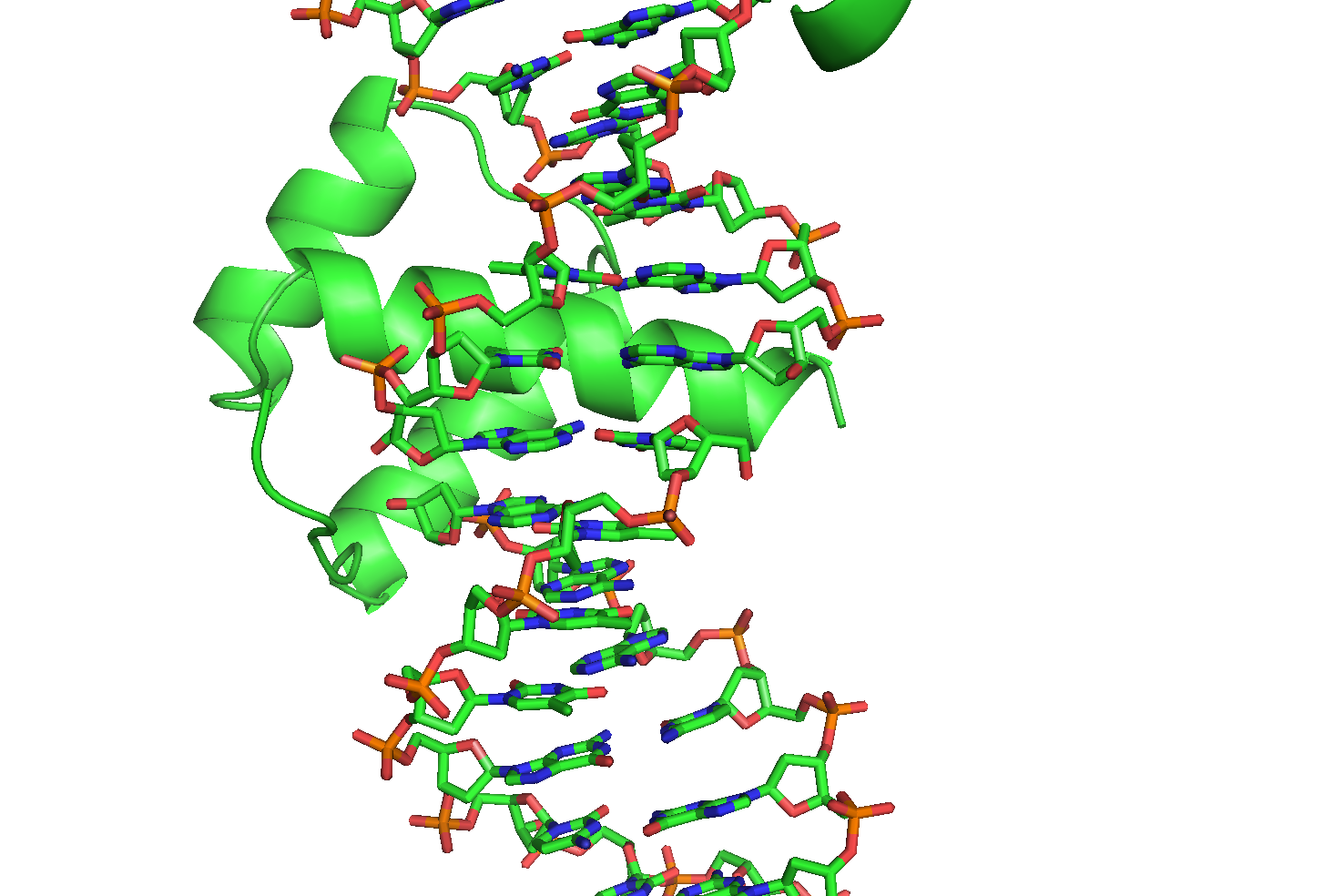
Рисунок 12 Взаимодействие липких концов разных фрагментов ДНК
Осталось понять, почему торцевой гомеодомен взаимодействует с ДНК как-раз в зоне разрыва. Оказывается, что 3'-конец первого фрагмента в зоне контакта
оканчивается на ТАА, а следующий фрагмент продолжает его своим липким концом буквами ТТТ. Образуется мотив ТААТТ, который идеально подходит для посадки
гомеодомена.
Выводы
Ассиметрическая единица не всегда несет полную информацию, необходимую для "биологического" объяснения структуры. Чтобы не потерять эту информацию, необходимо
рассматривать все окружение молекулы в попытке восстановить правильную структуру.