Исследование структуры тРНК
Из базы был получен документ c PDB-кодом 1il2. Он содержит информацию о структуре белок-нуклеинового комплекса : нуклеиновая кислота – аспартил-тРНК и белок – аспартил-тРНК-синтетаза.
Каждый из компонентов представлен двумя цепями: белок – A и B, РНК – C и D. Аминоацил-тРНК-синтетазы функционируют в клетке как димеры, и каждая связывает в активном центре по молекуле тРНК, соответственно, в файле видим 2 молекулы тРНК. Но димер аминоацил-тРНК-синтетазы может связывать в своих активных центрах разные изоакцепторные аспартил-тРНК, однако из поля SEQRES pdb-файла узнаем, что последовательности цепей C и D одинаковы, поэтому выберем для удобства дальнейшего изучения цепьC.
Заметим, что составляющие комплекса получены из разных организмов (белок – из E.coli, РНК – из Sacchromyces cereviciae), что позволяет предположить, что кристалл для рентгеноструктурного анализа был получен в лабораторных условиях "смешением" выделенных ранее структур.
Исследуемая тРНК состоит из 75-ти остатков. Ниже приведена ее нуклеотидная последовательность:
| U | C | C | G | U | G | A | U | A | G | U | U | PSU |
| A | A | H2U | G | G | H2U | C | A | G | A | A | U | G |
| G | G | C | G | C | PSU | U | G | U | C | 1MG | C | G |
| U | G | C | C | A | G | A | U | 5MC | G | G | G | G |
| 5MU | PSU | C | A | A | U | U | C | C | C | C | G | U |
| C | G | C | G | G | A | G | C | C | A |
Как видим, 8 из 75-ти остатков являются тем или иным образом модифицированными. Среди них следующие "нетипичные" нуклеотиды:
- PSU
- Псевдоуридин-5'-монофосфат
- H2U
- 5,6-Дигидроуридин-5'-монофосфат
- 1MG
- 1N-метилгуанозин-5'-монофосфат
- 5MC
- 5-метилцитидин-5'-монофосфат
- 5MU
- 5-метилуридин-5'-монофосфат
Примечание: Для краткости будем называть нуклеозиды (дигидроуридин), имея в виду соответствующие нуклеотиды (5,6-дигидроуридин-5'-монофосфат).
Чтобы изучить нумерацию остатков тРНК, рассмотрим файл, полученный командой "save pdb"(с одной только цепью C). Используем возможности команды grep и найдем в этом файле все строки, содержащие "O1P" и получим список. Отметим:
- Нумерация начинается с 901 остатка, оканчивается на 976
- Номер 947 отсутствует
- Нет "лишних" номеров (вставок, "insertion codes")
Чтобы проанализировать вторичную структуру аспартил-тРНК, воспользуемся программой find_pair. Выполнив
find_pair -t 1IL2C.PDB pairs.txtполучаем в файле pairs.txt информацию о комплементарных парах, образующихся между основаниями аспартил-тРНК и, соответственно, о двуцепочечных участках вторичной структуры последней.Find_pair выдает информацию о водородных связях.
На основании результатов программы выделим 7 спиралей вторичной стуктуры. Далее, с помощью скрипта:
background white define helix1 901-907,966-972 define helix2 938-944, 926-932 define helix3 949-954, 958-965 define helix4 910-913, 922-925 select all wireframe off backbone 130 select helix1 color yellow select helix2 color orange select not (helix1, helix2, helix3, helix4) color gray select A976.p, U901.p cpk 80 label %r%nПолучим следующее изображение:
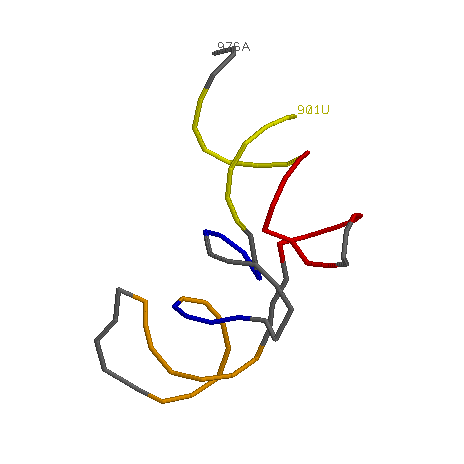 ,
,
Спирали третичной структуры
Таких участков три, причем один из них состоит всего из одной парыОбратимся подробнее к результатам поиска дуплексов. Нуклеотиды, участвующие в их образовании отмечены разными цветами (соответственно спирали, в составе которой они находятся):
- Спираль 1 (длина - 14 нуклеотидов) – желтый цвет
- Спираль 2 (длина - 15 нуклеотидов) – светло-оранжевый цвет
- Спираль 3 (длина - 1 нуклеотид) – коричневый цвет
.
| UCCGUA U A GUU PSU AA H2U G GH2UC H2UCAGAAUGGGCGCPSU UGUC 1MG CGUGCCAGAU 5MC GGGG 5MU PSU C A A U UCCCCGUCGCGGA GCCA |
Судя по раскраске последовательности можно понять, что преобладающая часть нуклеотидов аспартил-тРНК образует дуплексы (в их состав входят 60 из 75 остатков).
Такие же результаты выдает скрипт col_helices.scr, который находился в выдаче программы find_pair.
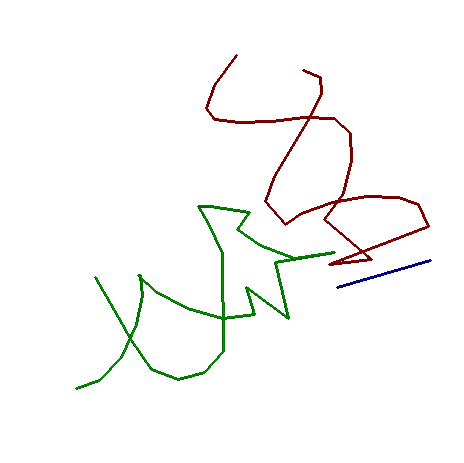
Для создания изображения был использован файл 1il2C.pdb, полученный при выполнении предыдущих заданий с помощью команды RasMol save pdb. Файл содержит структуру только изучаемой цепи C аспартил-тРНК.
background white define helix1 901-907, 949-955, 961-972, 917, 958 define helix2 938-944, 908, 910-916, 921-932, 948, 959, 946 define helix3 918, 956 select all wireframe off backbone 130 select helix1 color yellow select helix2 color orange select helix3 color red select not (helix1, helix2, helix3) color gray select A976.p, U901.p cpk 80 label %r%nПолучаем изображение нашей тРНК в остовной модели, с раскрашиванием по спиралям третичной структуры.
 Желтым цветом
отображены нуклеотиды (точнее, соответствующие им части сахарофосфатного остова), принимающие участие в формировании первой
спирали; оранжевым – второй, красным – два нуклеотида третьей. Серым показаны немногочисленные
остатки одноцепочечных участков.
Желтым цветом
отображены нуклеотиды (точнее, соответствующие им части сахарофосфатного остова), принимающие участие в формировании первой
спирали; оранжевым – второй, красным – два нуклеотида третьей. Серым показаны немногочисленные
остатки одноцепочечных участков.
Рассмотрим 945-ый гуанин и 946-ой аденин(отмечены зеленым цветом)
 В данном
случае
ожидаемо увидеть внеспиральный стекинг.
Также отметим неканоническое взаимодействие G-U
В данном
случае
ожидаемо увидеть внеспиральный стекинг.
Также отметим неканоническое взаимодействие G-U
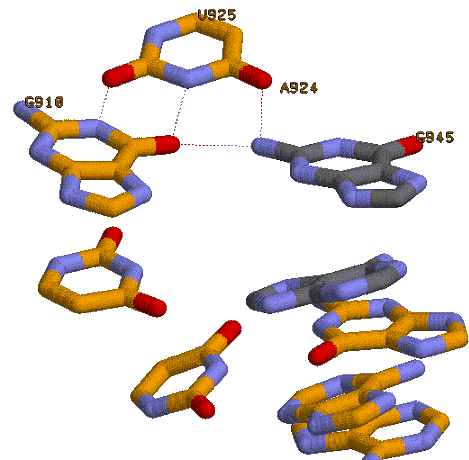 На рисунке также видно связывание данной пары еще и с третьим нуклеотидом - гуанином 945, не принадлежащим спирали 2, ведущее к образованию
триплета. Данный нуклеотид показан на рисунке иным цветом. Ниже взаимодействующих с ним остатков спирали 2
показаны дополнительно остатки этой структуры. Интересно, что с внеспиральным гуанином взаимодействуют оба
основания пары (у атома гуанина два донорных атома водорода, а оба подходящих к нему близко атома – акцепторы).
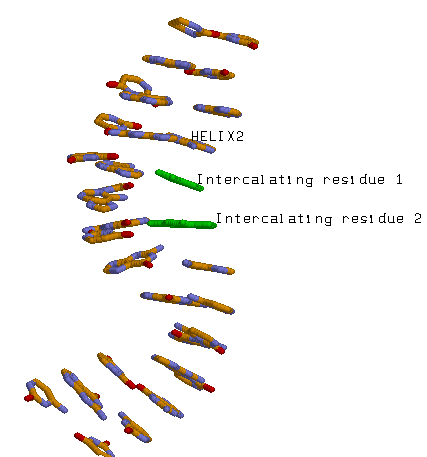
Спирали тРНК больше похожи на А-форму ДНК, так как обладают свойством двуцепочечного стекинга(плоскости оснований одной цепи дуплекса могут взаимодействовать с плоскостями не только той же, но и второй цепи (что невозможно в форме B)). Рассмотрим первую спираль.
При рассмотрении в RasMol заметим, что спираль имеет 11 нуклеотидов на виток, с торца можно увидеть "канал", поэтому сделаем вывод о том, что форма - А.
Проверим выводы с помощью программы analyze
Файл helix1.out. Программа подтверждает, что это A-форма.
Таким образом, сделанные выше с помощью RasMol
выводы верны.
На рисунке также видно связывание данной пары еще и с третьим нуклеотидом - гуанином 945, не принадлежащим спирали 2, ведущее к образованию
триплета. Данный нуклеотид показан на рисунке иным цветом. Ниже взаимодействующих с ним остатков спирали 2
показаны дополнительно остатки этой структуры. Интересно, что с внеспиральным гуанином взаимодействуют оба
основания пары (у атома гуанина два донорных атома водорода, а оба подходящих к нему близко атома – акцепторы).
Спирали тРНК больше похожи на А-форму ДНК, так как обладают свойством двуцепочечного стекинга(плоскости оснований одной цепи дуплекса могут взаимодействовать с плоскостями не только той же, но и второй цепи (что невозможно в форме B)). Рассмотрим первую спираль.
При рассмотрении в RasMol заметим, что спираль имеет 11 нуклеотидов на виток, с торца можно увидеть "канал", поэтому сделаем вывод о том, что форма - А.
Проверим выводы с помощью программы analyze
Файл helix1.out. Программа подтверждает, что это A-форма.
Таким образом, сделанные выше с помощью RasMol
выводы верны.
При рассмотрении второй и третьей спиралей возникают трудности, так как вторая очень изломана и имеет нерегулярную структуру, а третья состоит лишь из одной пары нуклеотидов. Но судя по тому, что во второй спирали с торца удается увидеть небольшое отверстие, можно предположить, образно говоря, что и ей A-форма не чужда.
Для работы с программой einverted из текста pdb-файла была получена и сохранена в fasta-формате последовательность аспартил-тРНК. C минорными нуклеотидами можно было поступить двояко: заменить их обозначения в последовательности на
- N – любой нуклеотид
- обозначение близкого канонического нуклеотида (PSU,H2U → U; 2MC → C и т.п.)
Попробовали сделать и так, и так. При запуске программы einverted нам задавались вопросы. При этом мы поменяли только один из параметров – "Minimum score threshold", его пришлось сделать очень маленьким (5 вместо 50), только в таком случае выдавались какие-то результаты( Участки очень короткие, веса маленькие, поэтому и порог низкий) В результате для двух рассматриваемых случаев получили разные результаты. Если мы создавали неопределенность, записывая минорные нуклеотиды как N, программа "увидела" два инвертированных участка, если же использовали канонические аналоги – только один. Участки короткие и содержат в обоих случаях только канонические нуклеотиды (никаких остатков N). Значит, наличие или отсутствие неопределенности влияет каким-то образом на поиск инвертированных повторов, однако точно определить данное влияние сложно. Вероятно, наличие позиций, которые может занимать любой нуклеотид, дает больше степеней свободы в поиске, позволяет рассматривать большее разнообразие структур, поэтому выдается больше результатов.
В первом случае в одном из двух участков все основания комплементарны, в другом есть одна некомплементарная пара GT. Если же взять второй случай (где в последовательности неканонические нуклеотиды заменялись на обычные), то выдается только один участок (первый из вышеописанных, с полным спариванием).
Однако в любом случае выдается не более двух коротких участков инвертированных повторов (которые образуют спирали). Программа einverted способна показать нам только какой-то участок полной спирали, и то не всегда. Результатов поиска инвертированных повторов явно недостаточно для того, чтобы охарактеризовать вторичную структуру РНК, ведь она намного сложнее, чем совокупность таких повторов, образующих дуплексы, и одноцепочечных участков.
В директорию рабочей папки поместили файл с последовательностью тРНК в фаста-формате; минорные нуклеотиды заменили схожими нуклеотидами из стандартного набора. Для работы использовали такую команду:
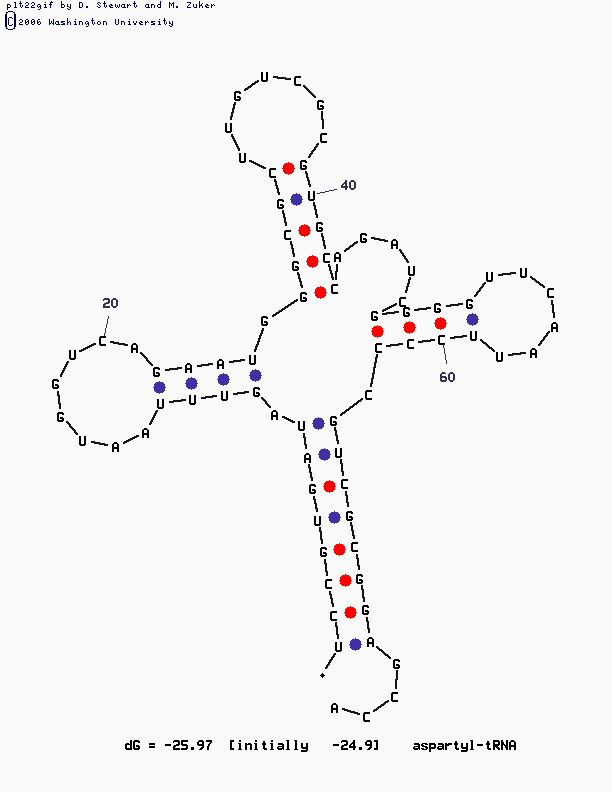
mfold SEQ=tRNA.fasta P=Xгде X – значение параметра P – число, означающее, на сколько процентов выдаваемое предсказание вторичной структуры может отличаться по своей вычисленной энергии от оптимального. В выдачах mfold необходимые нам файлы с расширением GIF, демонстрирующие изображение предсказанной структуры. Понятно, что чем больше значение P, тем больше степеней свободы и, соответственно, количество выдаваемых структур.
Наиболее удовлетворительной показалась третья по счету структура, полученная при втором обращении к mfold (со значением P, равным 15-ти). Все, кроме выбранной, либо не похожи на клеверный лист вовсе, либо не обладают одним из стеблей.