Вторичная структура тРНК и ДНК-белковые взаимодействия
Задание 1
Было проведено предсказание координат стеблей в структуре глутамил-тРНК Thermus thermophilus (PDB ID: 1n78) при помощи программы einverted из пакета EMBOSS и по алгоритму Зукера при помощи ViennaRNA (Рис. 1).
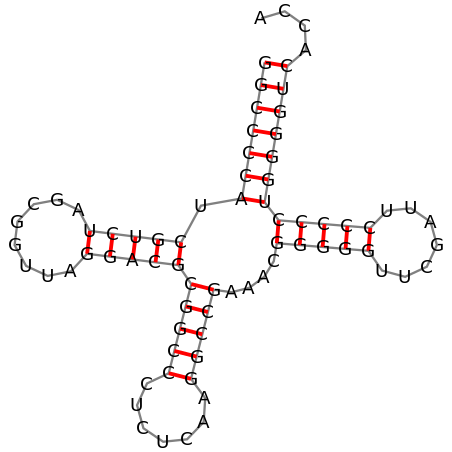
Сравнение результатов предсказания приведено в Таблице 1. Для программы einverted были использованы следующие параметры: -gap 12 -threshold 16 -match 4 -mismatch -9 -maxrepeat 20, вследствие чего (параметр maxrepeat) акцепторный стебель в полученном предсказании отсутствовал.
| Участок структуры | findpair | einverted | ViennaRNA |
|---|---|---|---|
| Акцепторный стебель | 1-7 (66-72) | - | 1-7 (66-72) |
| T-стебель | 26-32 (38-44) | 27-31 (39-43) | 27-31 (39-43) |
| D-стебель | 10-14 (21-25) | 9-12 (23-26) | 9-13 (22-26) |
| Антикодоновый стебель | 49-53 (61-65) | 49-53 (61-65) | 49-53 (61-65) |
| Общее число канонических пар | 20 | 14 | 20 |
Можно заметить, что предсказания einverted и ViennaRNA на трех из четырех стеблей совпадают и для T- и D-стеблей несколько отличаются от предсказания, полученного при помощи findpair. Однако, общее количество предсказанных канонических пар совпадает для результатов findpair и ViennaRNA, а результат einverted отличается от них только из-за отсутствия акцепторного стебля в предсказании.
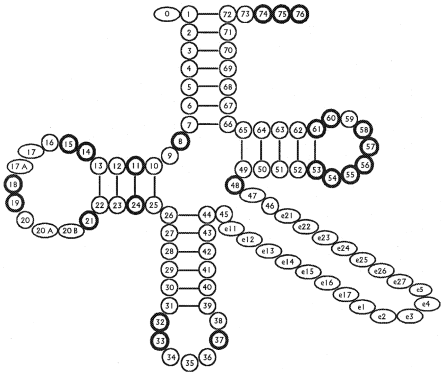
При сравнении результатов предсказания было необходимо учитывать тот факт, что нумерация нуклеотидов в тРНК не сквозная, а подчиняется правилам, основанным на выравниваниях и сравнениях структур различных тРНК (Рис. 2). Так в рассматриваемой глутамил-тРНК бактерии Thermus thermophilus присутствует нуклеотид с номером 520A, но отсутствуют с номерами 517 и 547 (в файлах PDB, включающих одновременно структуру тРНК и АРСазы нумерация нуклеотидов тРНК начинается не с 1, а с 501).
Задание 2
Скрипт, определяющий необходимые множества атомов
Скрипт, выводящий изображение всей структуры
Рассматриваемая структура (PDB ID: 1I3J) описывает взаимодействие ДНК-связывающего домена интронной хоуминг-эндонуклеазы I-TevI с дуплексом ДНК. Данный домен содержит мотив "цинковый палец", протяженный участок, взаимодействующий с малой бороздкой, и мотив спираль-поворот-спираль (HTH) [2]. Для этой структуры были рассмотрены полярные и неполярные контакты ДНК-белок (Табл. 2).
| Контактирующие атомы ДНК | Полярные | Неполярные | Всего |
|---|---|---|---|
| Отстатки 2'-дезоксирибозы | 10 | 120 | 130 |
| Остатки фосфорной кислоты | 24 | 27 | 51 |
| Азотистые основания со стороны большой бороздки | 0 | 25 | 25 |
| Азотистые основания со стороны малой бороздки | 3 | 10 | 13 |
Также была построена карта ДНК-белковых контактов при помощи nucplot (Рис. 3).
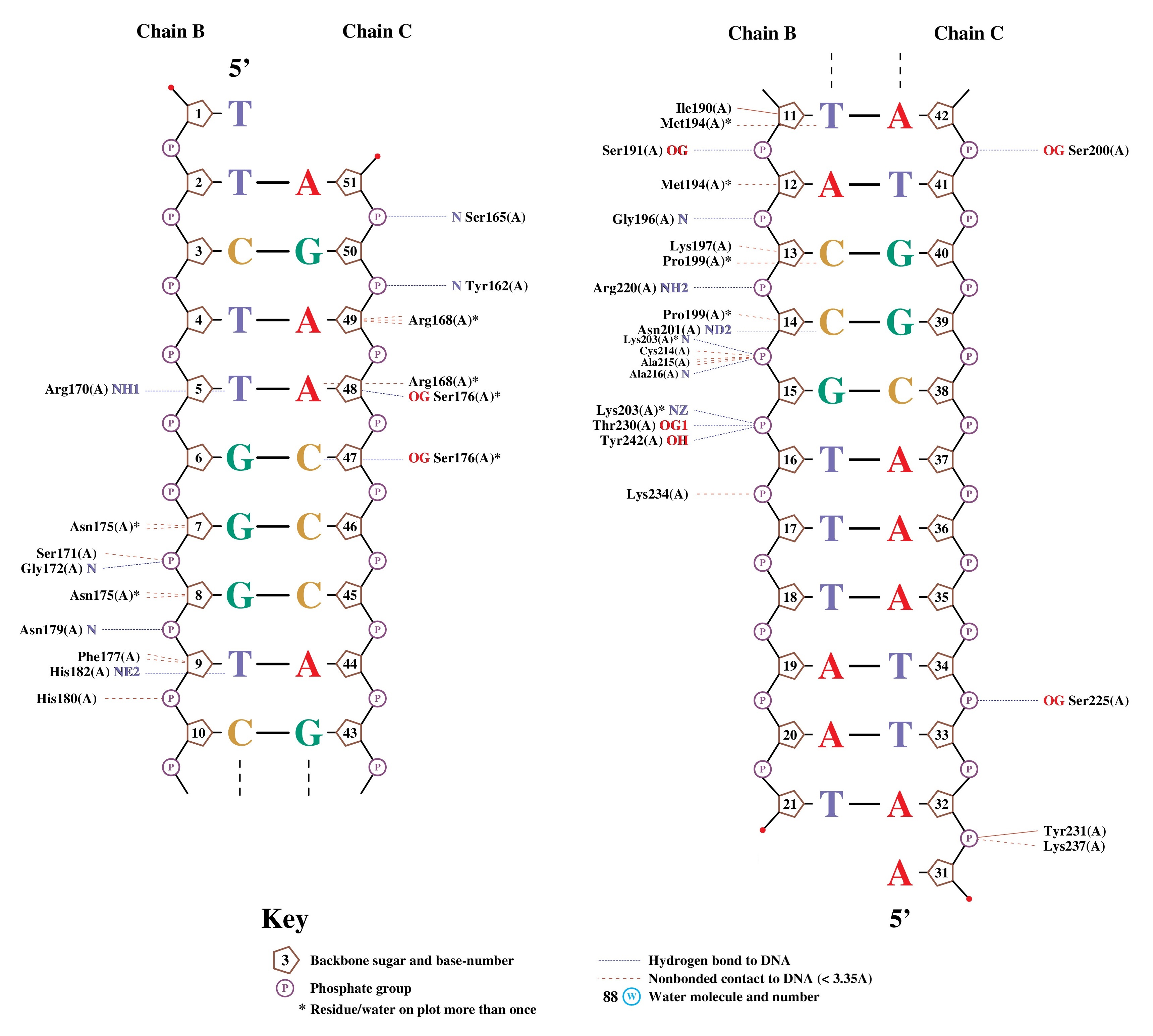
Согласно карте nucplot наибольшее число контактов с ДНК образует аминокислотный остаток N175, взаимодействующий с нуклеотидными остатками G7 и G8 через малую бороздку (Рис. 4).
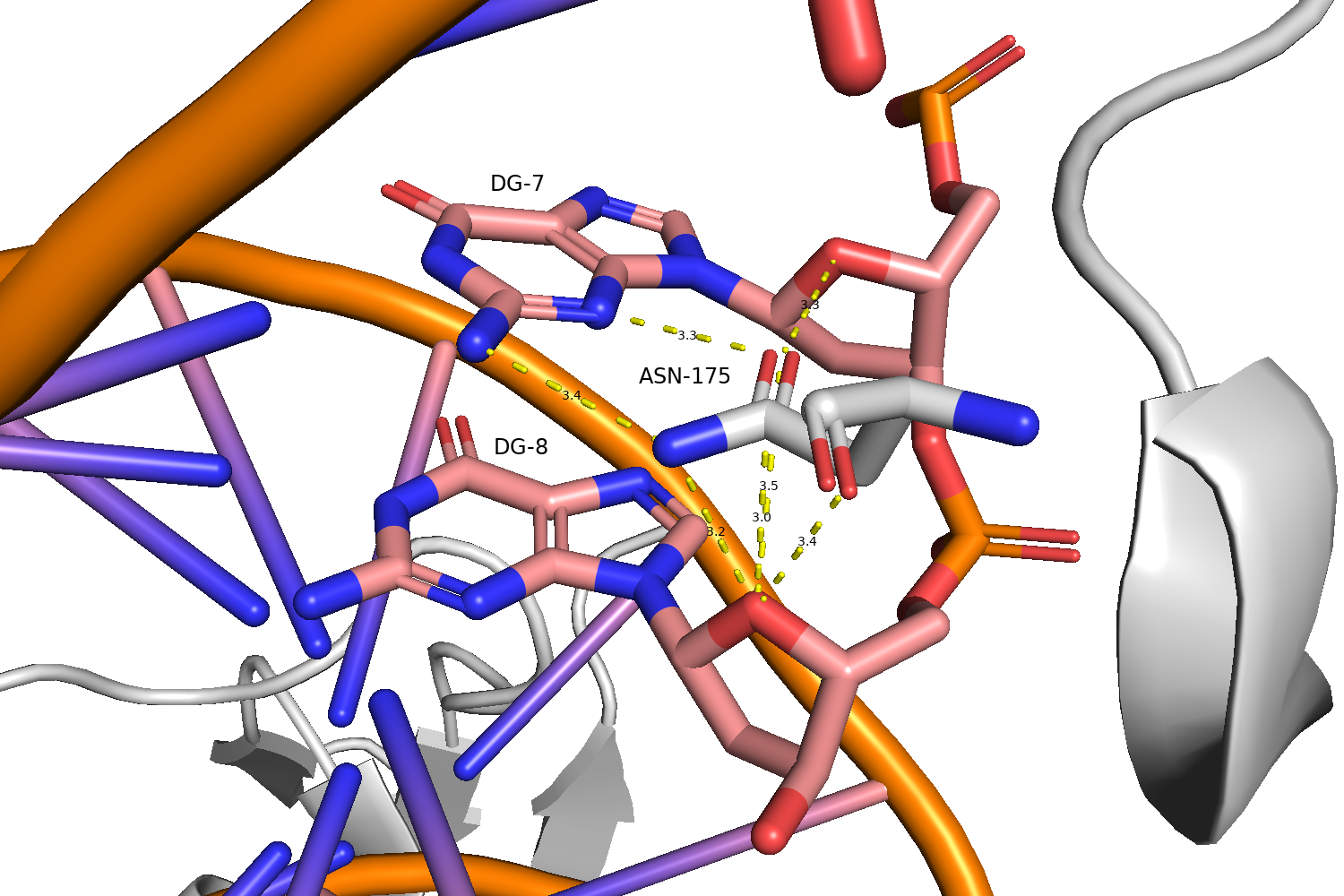
Стоит отметить, что nucplot определяет неполярные контакты только в том случае, когда атомы находятся на расстоянии менее 3.35 Å друг от друга, вследствие чего часть неполярных контактов теряется, поэтому для поиска наиболее важных для распознавания последовательности взаимодействий использовались контакты определенные ранее в PyMol вручную.
В представленной структуре обнаруживается малое количество полярных контактов, причем большая их часть связана с сахарофосфатным остовом (Рис. 5), остальные - с малой бороздкой ДНК (Рис. 6). Это, вероятно, является причиной толерантности нуклеазы I-TevI к последовательности [2] и отличает ее от представителей наиболее многочисленного семейства хоуминговых нуклеаз LAGLIDADG (I-TevI относится к семейству GIY-YIG), для которых хаарктерно связывание преимущественно с большой бороздкой.
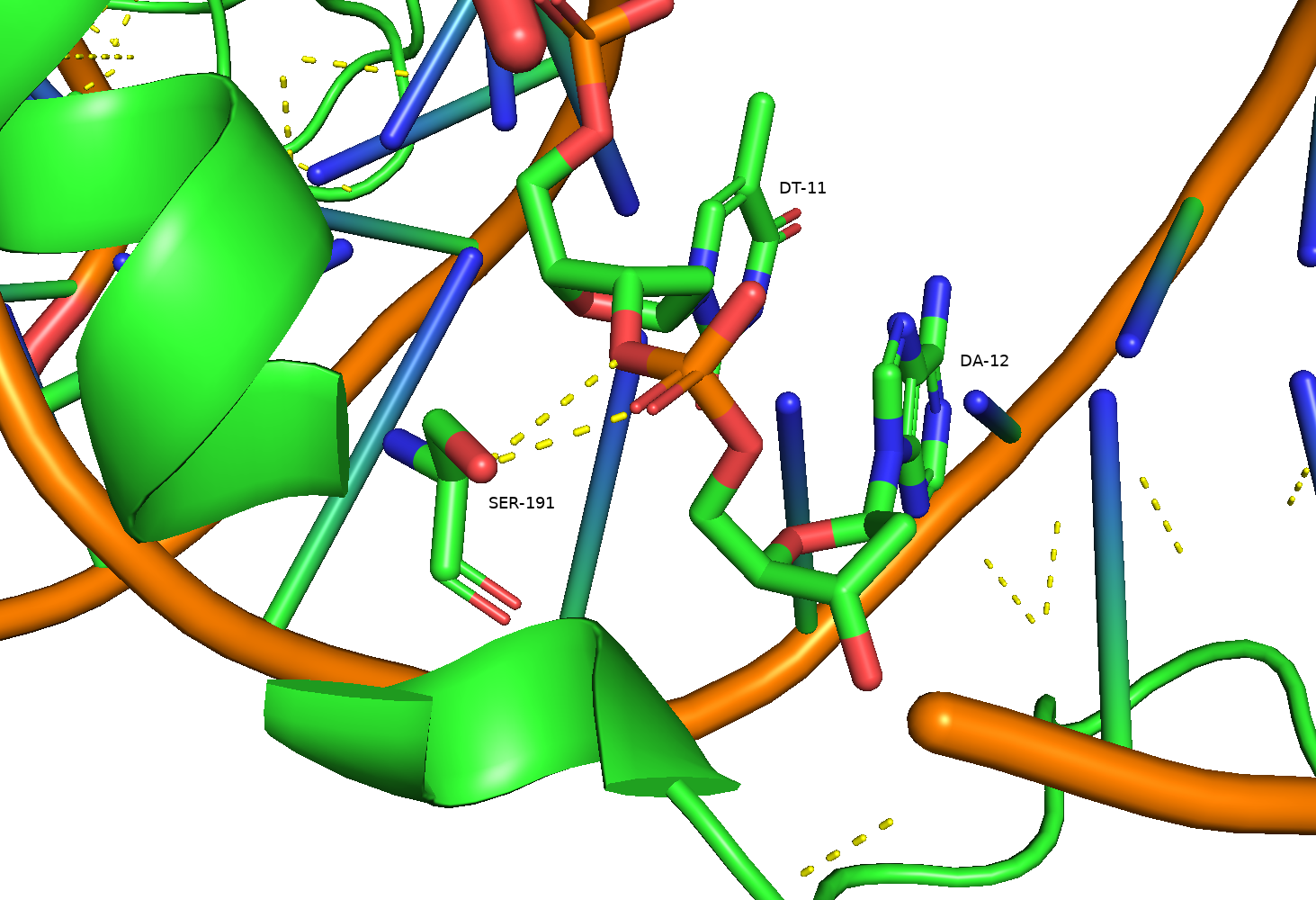

Однако аминокислотные остатки мотива HTH взаимодействуют с ДНК посредством неполярных взаимодействий, связываясь с 5-метильными группами тиминовых азотистых оснований (Рис. 7), вследствие чего можно предположить, что эти взаимодействия отвечают за специфичность данного домена по отношению к AT-богатым участкам.

ИСТОЧНИКИ
Mathias Sprinzl, Carsten Horn, Melissa Brown, Anatoli Ioudovitch, Sergey Steinberg, Compilation of tRNA sequences and sequences of tRNA genes, Nucleic Acids Research, Volume 26, Issue 1, 1 January 1998, Pages 148–153, https://doi.org/10.1093/nar/26.1.148
Roey PV, Belfort M, Derbyshire V. Homing Endonuclease I-TevI: An Atypical Zinc Finger with a Novel Function. In: Madame Curie Bioscience Database [Internet]. Austin (TX): Landes Bioscience; 2000-2013. Available from: https://www.ncbi.nlm.nih.gov/books/NBK6328/