Поиск и реконструкция филогении гомологов АТФ-азы ClpX Escherichia coli
ClpX - представитель суперсемейства AAA+ АТФаз, формирующий мультисубъединичный комплекс с протеазой ClpP (два гексамерных кольца из ClpX образуют комплекс с тетрадекамером из двух гептамерных колец ClpP), играющий важную роль в жизнедеятельности бактериальной клетки. Данный комплекс осуществляет деградацию белков, при этом ClpX является шапероном, отвечающим за их разворачивание (анфолдазой). Благодаря своей способности АТФ-зависимо протягивать через себя аминокислотную последовательность белка, ClpX находит применение в разрабатываемых методиках нанопорового секвенирования белков [1].
Поиск гомологов ClpX E.coli
Поиск гомологов ClpX Escherichia coli был осуществлен при помощи blastp по протеомам бактерий Agrobacterium fabrum (AGRFC), Aromatoleum aromaticum (AROAE), Bordetella pertussis (BORPE), Burkholderia mallei (BURMA), Haemophilus influenzae (HAEIN), Neisseria meningitidis (NEIMA), Polynucleobacter asymbioticus (POLAQ), Thiobacillus denitrificans (THIDA). Из данных протеомов была сформирована база данных для blastp. также для выбранных бактерий было построено предполагаемое дерево по таксономии (Рис. 1).
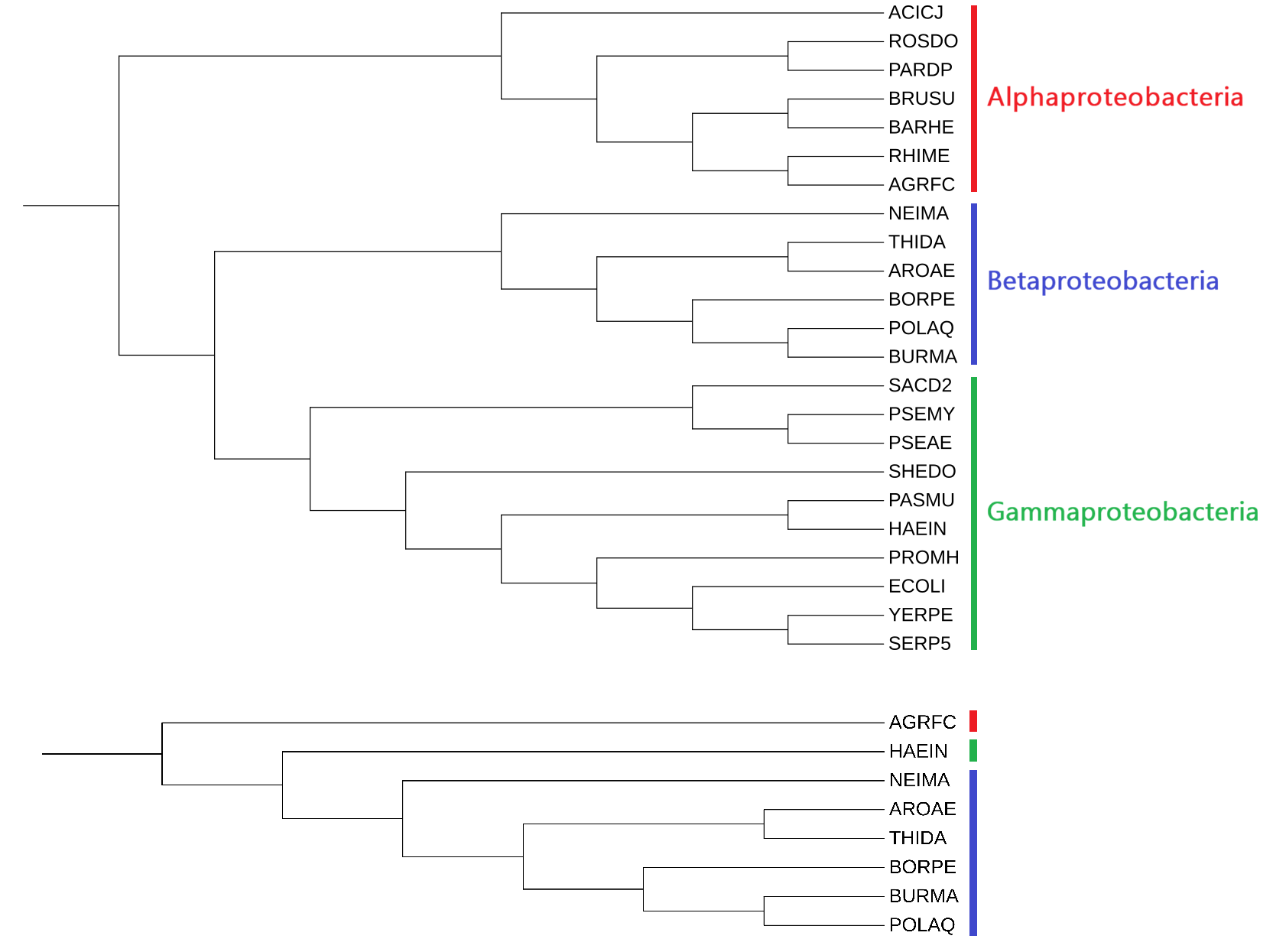
Для поиска достоверных гомологов был выбран порог e-value 10-5, так как при меньшем пороге (10-4) среди находок оказываются такие белки как FtsH, для которых покрытие выравниванием составляет менее 40%.
blastp -query ClpX.txt -db proteomes -out clpx_res.tsv -outfmt 6 -evalue 1e-5
Среди 14 находок имелись 8 белков ClpX и 6 HslU (также AAA+ АТФаза-анфолдаза, образующая комплекс с протеазой HslV), что интересно, для всех последовательностей HslU имелось 2 хита с последовательностью запроса. Файл, содержащий последовательности находок был загружен из выдачи онлайн-инструмента BLAST.
Реконструкция филогении гомологов ClpX E.coli
Полученные на предыдущем этапе последовательности достоверных гомологов ClpX E.coli были выровнены при помощи алгоритма MAFFT:
mafft ClpX_homologues_noFTSH.fasta > ClpX_noFTSH_aligned.fasta
Затем на основании полученного выравнивания было построено филогенетическое дерево при помощи программы iqtree с настройками по умолчанию и ультрабыстрой бутстреп-проверкой из 1000 реплик:
iqtree -s ClpX_noFTSH_aligned.fasta -bb 1000
Как можно видеть, полученное дерево (Рис. 2) при укоренении в среднюю точку разбивается на 2 поддерева, содержащих последовательности ClpX или HslU.
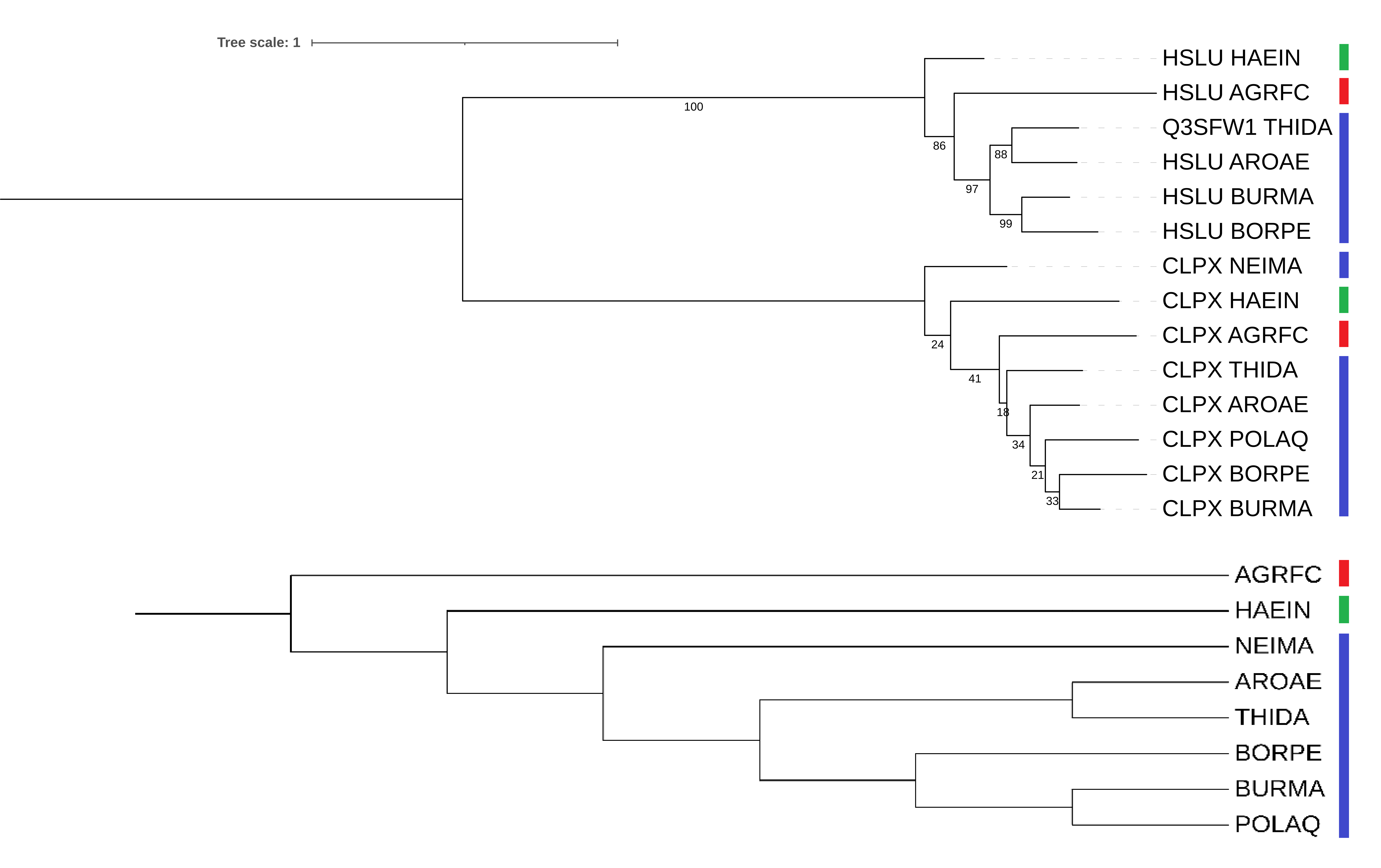
Внутри клады ClpX базальной ко всем остальным оказывается бактерия NEIMA, которая по предполагаемой правильной филогении должна быть базальной среди выбранных бета-протеобактерий, но не среди всех бактерий. Также несмотря на то, что BURMA, BORPE и POLAQ образуют монофилетическую группу, ближайшими родственниками оказываются BORPE и BURMA, но не BURMA и POLAQ. Монофилетическая группа из AROAE и THIDA отсутствует, можно также отметить низкую поддержку всех ветвей в поддереве ClpX.
Внутри группы HslU поддержка ветвей оказывается выше, и присутствуют монофилетические группы из BURMA и BORPE, а также THIDA и AROAE. Стоит отметить, что HslU был найден не у всех бактерий, что вероятно, связано с его меньшей распространенностью по сравнению с ClpX, которая в свою очередь может быть связана с утратой данного белка некоторыми группами в ходе эволюции или, наоборот, приобретением его некоторыми группами посредством ГПГ.
Помимо прочего, видно, что положения единственных альфапротеобактерии (AGRFC) и гаммапротеобактерии (HAEIN) в обоих поддеревьях оказались верными лишь отчасти, они действительно базальны по отношению к бетапротеобактериям (или к их большей части в случае ClpX), но сестринской группой для них является AGRFC, но не HAEIN.
Исходя из того, что белки ClpX и HslU кластеризуются в отдельные группы (Рис. 3), можно считать ортологами любую пару, содержащую один белок, происходящий из разных бактерий, например, ClpX Burkholderia mallei (BURMA) и ClpX Bordetella pertussis (BORPE), ClpX Haemophilus influenzae (HAEIN) и ClpX Thiobacillus denitrificans (THIDA), HslU Agrobacterium fabrum и HslU Aromatoleum aromaticum (AROAE).
Паралогами же (точнее, ин-паралогами) можно считать любую пару разных белков, происходящих из одной бактерии, например, ClpX и HslU Burkholderia mallei (BURMA), ClpX и HslU Polynucleobacter asymbioticus (POLAQ), ClpX и HslU Thiobacillus denitrificans (THIDA).
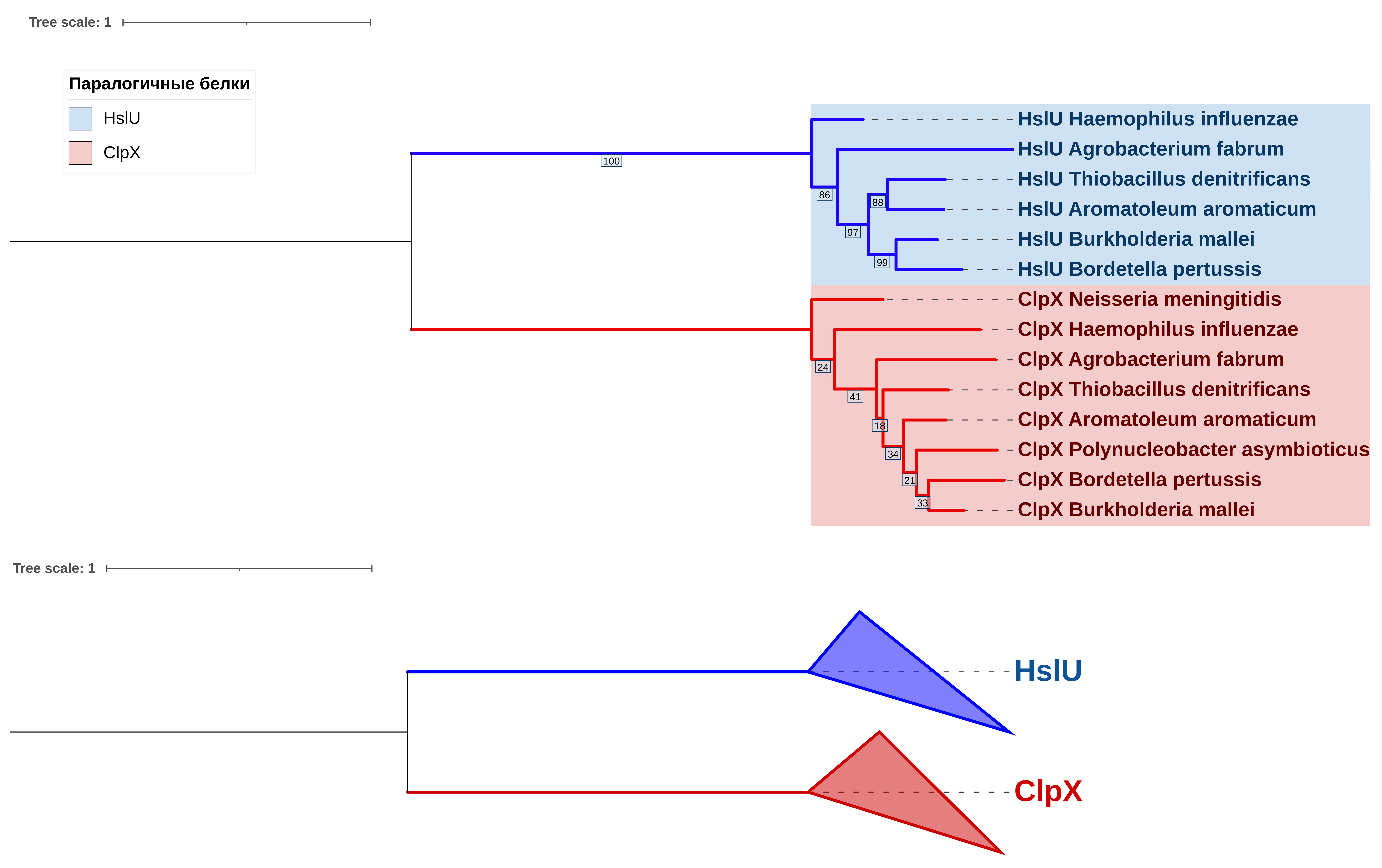
Как можно предположить исходя из топологии дерева, разделение ClpX и HslU произошло у предков рассматриваемых групп псевдомонадот. Также, поскольку ожидаемая топология по таксономии в поддеревьях оказалась искажена незначительно, вполне вероятно, что эти белки эволюционировали вертикально, несмотря на распространенность горизонтального переноса генов у прокариот.
ЛИТЕРАТУРА И ИСТОЧНИКИ
Motone, K., Kontogiorgos-Heintz, D., Wee, J. et al. Multi-pass, single-molecule nanopore reading of long protein strands. Nature 633, 662–669 (2024). https://doi.org/10.1038/s41586-024-07935-7