 8 (926) 907 94 08
8 (926) 907 94 08
Всё на свете является чудом!
A- и В- формы ДНК. Структура РНК
При помощи программы Putty мы подсоединились к серверу kodomo.cmm.msu.ru, после чего перешли в рабочую директорию командой cd Term3/Practice2, предварительно создав эту директорию командой mkdir Practice2.
Для указания пути к 3DNA мы ввели команды: export PATH=${PATH}:/home/preps/golovin/progs/X3DNA/bin и
export X3DNA=/home/preps/golovin/progs/X3DNA.
Далее с помощью программы fiber пакета 3DNA командой fiber -* gatc-*.pdb, где "*" соответствует необходимой форме (a, b и z), мы построили, а затем и сохранили три формы дуплекса ДНК: А-форму, В-форму и Z-форму.
Задание 2. Работа со структурами нуклеиновых кислот средствами Rasmol
Упражнение #1 (выделение разных атомов и химических группировок, используя предопределенные множества RasMol)
Применяя скрипты последовательно для каждой формы дуплекса ДНК, мы получаем различные изображения:
1. сахарофосфатный остов ДНК
background white
zoom 100
restrict none
select backbone
wireframe 60
rotate y 90
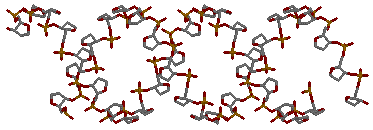 →
для gatc-a
→
для gatc-a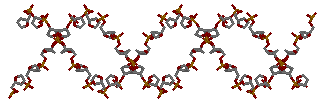 →
для gatc-b
→
для gatc-b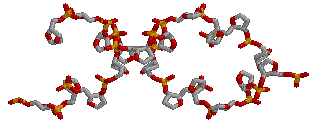 →
для gatc-z
→
для gatc-z2. все нуклеотиды
background white
zoom 100
restrict none
select A
color red
wireframe 60
select G
color blue
wireframe 60
select T
color yellow
wireframe 60
select C
color green
wireframe 60
rotate y 90
 → для
gtac-a
→ для
gtac-a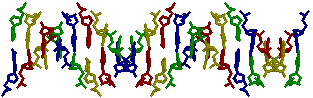 → для
gtac-b
→ для
gtac-b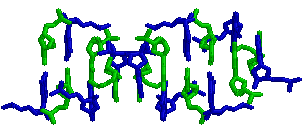 → для
gtac-z (как мы видим, аденина и тимина в
z-форме нет)
→ для
gtac-z (как мы видим, аденина и тимина в
z-форме нет)3. все аденины
background white
zoom 100
restrict none
select backbone
color orange
backbone 40
select A
color red
wireframe 60
rotate y 90
 → для
gtac-a
→ для
gtac-a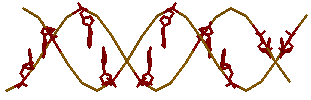 → для
gtac-b
→ для
gtac-b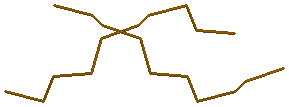 → для
gtac-z (ещё раз убеждаемся, что аденин в
z-форме отсутствует)
→ для
gtac-z (ещё раз убеждаемся, что аденин в
z-форме отсутствует)4.1 атом N7 во всех гуанинах
background white
zoom 100
restrict none
select backbone
color grey
backbone 40
select G
color blue
spacefill 50
wireframe 60
select G and *.N7
spacefill 150
color yellow
rotate y 90
 → для gtac-a
→ для gtac-a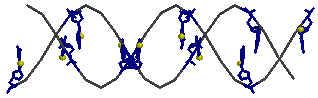 → для
gtac-b
→ для
gtac-b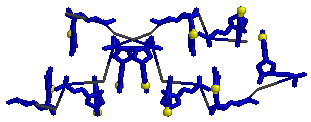 → для
gtac-z
→ для
gtac-z4.2 атом N7 в первом по последовательности гуанине
background white
zoom 100
restrict none
select backbone
color grey
backbone 40
select G
color blue
spacefill 50
wireframe 60
select G1 and *.N7
spacefill 150
color yellow
rotate y 90
 → для gtac-a
→ для gtac-a → для
gtac-b
→ для
gtac-b → для
gtac-z
→ для
gtac-zУпражнение #2 (получение файлов PDB)
На сайте PDB мы нашли документы, соответствующие заданным идентификаторам PDB, и сохранили их в формате pdb: 1QTQ.pdb и 1LRR.pdb.
Упражнение #3 (проверка заданных структур ДНК и РНК на наличие разрывов)
При проверке заданных структур 1QTQ и 1LRR разрывов обнаружено не было.
1QTQ и 1LRR в проволочной модели:
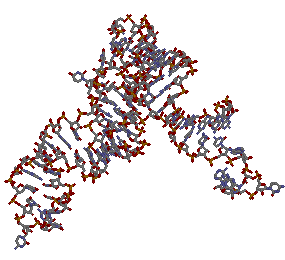 → RNA (1QTQ)
→ RNA (1QTQ)
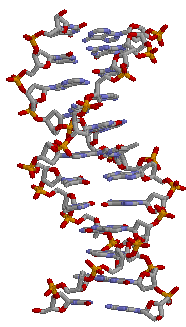 → DNA
(1LRR)
→ DNA
(1LRR)В завершении сохранили координаты атомов только ДНК и РНК в отдельных файлах.
Задание 3. Сравнение структур 3-х форм ДНК с помощью средств RasMol
Упражнение #1 (работа с большой и малой бороздками)
В Rasmol открыли файл gatc-b.pdb (B-форма).
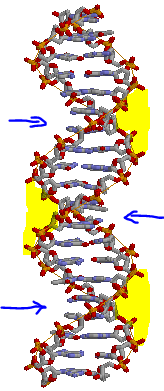
Визуально определили и отметили большую (отмечена синей стрелочкой)
и малую (выделена жёлтым цветом) бороздки.
На рисунке видно, что большая бороздка явно глубже малой.
Затем мы выбрали заданное азотистое основание (тимин) в произвольном месте структуры (T15 в цепи A). После чего выделили его и поработали с простейшими командами, чтобы лучше увидеть расположение атомов.
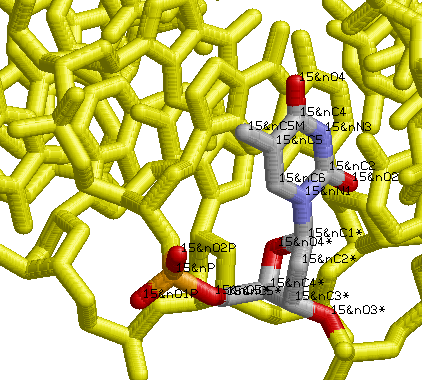
Определили, что:
○ в сторону большой бороздки обращены атомы
- T15.C4, T15.C5, T15.C5M, T15.C6 и T15.
○ в сторону малой бороздки обращены атомы
- T15.C2, T15.O2, T15.N3
○ примерно посередине находится атом T15.N1 (несмотря на небольшое сдвижение в сторону малой бороздки)
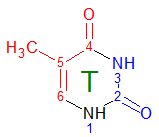
С помощью ChemSketch получили изображение нашего основания
и определённым образом раскрасили атомы, смотрящие в сторону большой и малой бороздок.
То же проделали с формами "A" и "Z".
Для формы A: в сторону большой бороздки в этой форме обращены все основания, и тимин в том числе, но в сторону малой бороздки немного повёрнуты атомы: T15.C4, T15.C5, T15.C5M, T15.C6 и T15.O4 (лишь повёрнуты, не обращены).
Для формы B: к сожалению, в этой форме не нашлось тимина, так как в ней представлены только цитозин и гуанин, поэтому взяли другой пиримидин вместо тимина - цитозин. При его изучении увидели, что в сторону большой бороздки обращены атомы C6.C2, C6.N3 и С6.O2, а в сторону малой бороздки - атомы C6.C5 и C6.C6. Примерно посередине находятся атомы С6.N1, С6.C4 и С6.N4.
Упражнение #2 (сравнение основных спиральных параметров разных форм ДНК)
| А-форма | В-форма | Z-форма | |
|---|---|---|---|
| Тип спирали | Правая | Правая | Левая |
| Шаг спирали (Å) | 28.03 | 33.75 | 43.50 |
| Число оснований на виток | 11 | 10 | 12 |
| Ширина большой бороздки (Å) | 16.81 (по аденину) | 17.21 (по гуанину) | 7.20 (по гуанину) |
| Ширина малой бороздки (Å) | 7.98 (по аденину) | 11.69 (по аденину) | 18.30 (по цитозину) |
Упражнение #3 (сравнение торсионных углов в структурах А- и В-форм)
| α (P-O5') | β (O5'-C5') | γ (C5'-C4') | δ (C4'-C3') | ε (C3'-O3') | ζ (O3'-P) | χ (C1'-N) | |
|---|---|---|---|---|---|---|---|
| А-ДНК | -51.68o | 174.79o | 41.73o | 79.05o или -161.74o | 100.35o | -75.10o | -94.92o |
| В-ДНК | -29.91o | 136.33o | 31.16o | 143.34o или -92.87o | 105.81o | -160.49o | -34.34o |
Полученные значения торсионных углов сильно отличаются от значений, приведенных в презентации, так как углы измерялись для тимина, а не для гуанина. Однако соотношение между углами А- и В-форм ДНК примерно соответствует такому соотношению для гуанина: углы α, β, γ, χ острее в В-форме, углы δ и ξ острее в А-форме.
Задание 4. Определение параметров структур нуклеиновых кислот с помощью программ пакета 3DNA
Упражнение #1 (определение торсионных углов нуклеотидов)
Командой find_pair -t gatc-*.pdb stdout | analyze (изменяя * на a, b, z), мы записали информацию о различных параметрах структур трёх форм ДНК в файлы gatc-*.out
Значения торсионных углов A-, B- и Z-форм выделили в отдельный файл.
Сравнивая значения соответствующих торсионных углов в структурах A-, B- и Z-форм ДНК, мы определили, что больше всего различаются значения углов:
○ A- и B-формы - δ и χ
○ A- и Z-формы - α
○ A- и Z-формы - α, χ и ξ
Командой remediator --old "XXXX.pdb" > "XXXX_old.pdb" сделали перевод pdb-файлов РНК и ДНК в старый формат, который приемлем для работы с программами пакета 3DNA.
Значения торсионных углов тРНК выделили в отдельный файл. При рассмотрении торсионных углов можно отметить сходство тРНК с A-формой ДНК.
Упражнение #2 (определение структуры водородных связей)
При помощи уже изученной программы find_pair из пакета 3DNA, мы определили возможные связи между азотистыми основаниями, получив на выходе файл с информацией.
Исходя из полученных данных выяснили:
○ акцепторный стебель состоит из участка 902-907 и комплементарного ему участка 966-971
○ T-стебель состоит из участка 949-953 и комплементарного ему участка 961-965
○ D-стебель состоит из участка 910-912 и комплементарного ему участка 923-925
○ антикодоновый стебель состоит из участка 926-933 и комплементарного ему участка 937-944
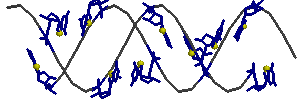
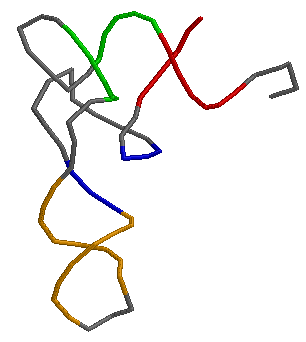
Используя RasMol, получили изображение остова исследуемой тРНК, где акцепторный стебель выделен красным, T-стебель - зелёным, D-стебель - синим, антикодоновый - оранжевым.
zap
load 1QTQ.PDB
background white
select all
color grey
backbone 100
wireframe off
select 902-907:B, 966-971:B
color red
select 949-953:B, 961-965:B
color green
select 910-912:B, 923-925:B
color blue
select 926-933:B, 937-944:B
color orange
В структуре отсутствуют тимидин (в T-петле) и дигидроуридины (в D-петле), но присутствует вариабельная петля (945-948).
Структуру стеблевых дуплексов поддерживают 19 канонических и 9 неканонических пар оснований.
Список неканонических пар:
1. U954 - A958
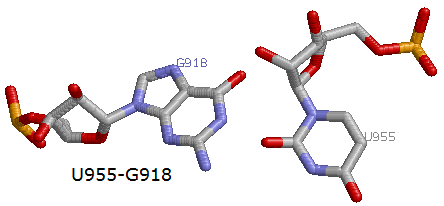
2. U955 - G918
3. A937 - U933
4. U938 - U932
5. C940 - G930
6. C944 - A926
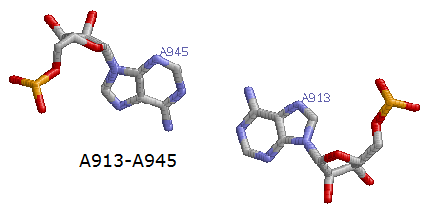
7. A913 - A945
8. A914 - U908
9. G915 - C948
Несмотря на наличие, казалось бы, канонических пар (G-C и A-U) среди неканонических, взаимодействия между этими основаниями не являются уотсон-криковскими из-за водородных связей между другими атомами.
Пример 2-х из 9-ти неканонических пар (2 и 7)
Исследуемая тРНК осуществляет транспорт глутаминовой кислоты. Она кодируется двумя триплетами: CAA и CAG. Следовательно возможные антикодоны - GUU и GUC. В антикодоновой петле исследуемой тРНК есть триплет GUC - это участок 934-936, представляющий собой антикодоновую петлю.
Что касается стабилизации третичной структуры тРНК, то присутствуют многочисленные дополнительные водородные связи, например, между основаниями D- и Т-петель есть две дополнительные водородные связи, одна из которых представляет собой каноническое взаимодействие (G919 - C956), другая - неканоническое (U955 - G918):
13 (0.004) B:.955_:[..U]Ux**+xG[..G]:.918_:B (0.013) x 28 (0.008) B:.919_:[..G]G-----C[..C]:.956_:B (0.003) +
Упражнение #3 (поиск стекинг-взаимодействий)
Рассмотрим несколько возможных вариантов:
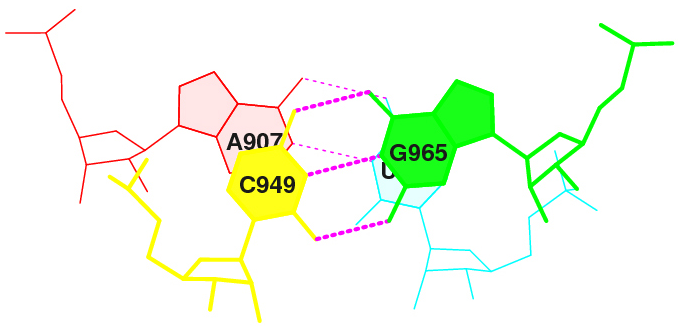
1) Стекинг-взаимодействия между основаниями конца акцепторного стебля и начала Т-стебля (это позиции 907-949 и им комплементарные 966-965):
6 (0.004) B:.907_:[..A]Ax----U[..U]:.966_:B (0.002) | 7 (0.002) B:.949_:[..C]C-----G[..G]:.965_:B (0.003) |Значения площадей перекрывания для этих пар такие:
step i1-i2 i1-j2 j1-i2 j1-j2 sum 6 AC/GU 2.22( 1.10) 0.00( 0.00) 0.00( 0.00) 5.96( 3.01) 8.18( 4.11)Как видим, перекрывание довольно значительное, и стекинг-взаимодействие вполне может присутствовать. Это иллюстрирует изображение, полученное с помощью команды stack2img:
2) Стекинг-взаимодействия между антикодоновым и D-стеблем (это позиции 944-910 и им комплементарные 926-925):
21 (0.005) B:.944_:[..C]Cx*---A[..A]:.926_:B (0.009) | 22 (0.005) B:.910_:[..G]G-----C[..C]:.925_:B (0.003) |
Значения площадей перекрывания для этих пар:
step i1-i2 i1-j2 j1-i2 j1-j2 sum 21 CG/CA 0.00( 0.00) 0.00( 0.00) 0.00( 0.00) 0.94( 0.28) 0.95( 0.28)
Как видим, перекрывание довольно небольшое, поэтому вероятность стекинг-взаимодействия чрезвычайно мала. Это подтверждает изображение:
| Главная | Об авторе | Учебные семестры | Проекты автора | Друзья | Ссылки партнеров | Extra | Контакты |
Mneff © 2011-2012