Описание тубулин-подобного белка CetZ2
Данный белок был выделен из археи Haloferax volcanii и представляет собой гомолог эукариотического тубулина и бактериального FtsZ [1]. Тубулин участвует в процессах направленного внутриклеточного транспорта, жгутикового движения и поддержания полярности субклеточных структур у эукариот [5]. FtsZ, найденный у большинства бактерий, формирует кольцевую структуру под мембраной при делении клетки. Данная кольцевая структура может самопроизвольно стягиваться, как показано в исследовании, проведённом на липосомах [6]. В исследовании [1] было установлено, что белок CetZ2 играет роль в контроле формы клетки археи, что в свою очередь важно для нормального её передвижения. В случае мутации Е212А, нарушающей функцию белка, были получены малоподвижные клетки с сильно неровными краями. Таким образом, CetZ2 вместе со своим близким гомологом CetZ1 являются перспективными белками для установления функциональной гомологии между цитоскелетными системами бактерий, архей и эукариот.
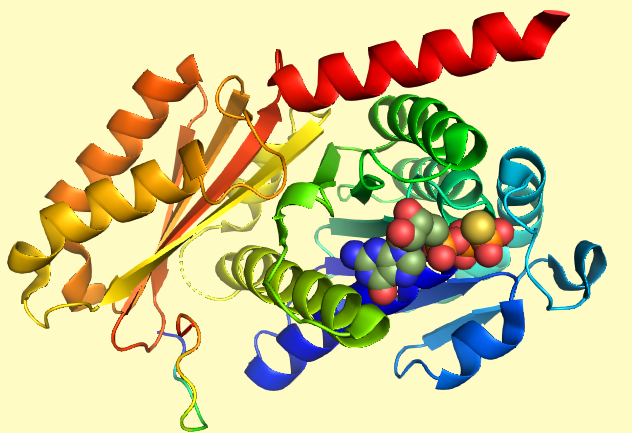
Изображение получено при помощи программы PyMOL
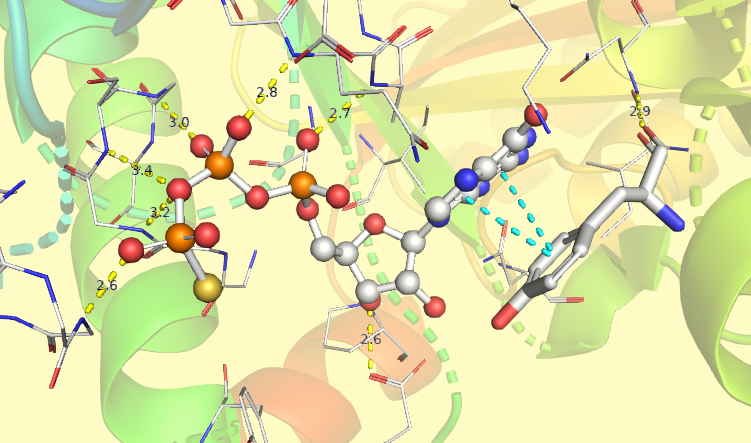
Изображение получено при помощи программы PyMOL
Как видно из поля GO текстовой версии страницы белка CetZ на UniProt [3], он имеет ГТФазный домен, подобный таковому у тубулина и FtsZ [4]. Анализируя структуру комплекса CetZ2 с негидролизуемым аналогом ГТФ гуанозиндифосфомонотиофосфатом [2] можно заметить, что основную роль в связывании субстрата играют водородные связи между остовными и амидными атомами азота белка с атомами кислорода фосфатных групп, а также параллельный стекинг между боковым радикалом тирозина и ароматической структурой гуанина, что проиллюстрировано на рисунке 2. Гидролиз ГТФ нужен для обеспечения самопроизвольной сборки белковых субъединиц в филаменты.
Haloferax volcanii - галофильная архея, живущая в тёплых горько-солёных водоёмах (Мёртвое море, Большое Солёное озеро, некоторые части Мирового океана). Аэроб, но способна к анаэробному образу жизни. В отличие от многих архей нормально культивируется в лаборатории, что делает её ценным модельным объектом [7]. Клеточной стенки не имеет, однако клетки окружены S-слоем - характерным для архей слоем одинаковых белков, заякоренных в мембране и экспонированных наружу [8]. Клетки уплощённо-цилиндрические, 1-3 мкм в диаметре, накапливают ликопин в мембранах, что придаёт им алую окраску [9].
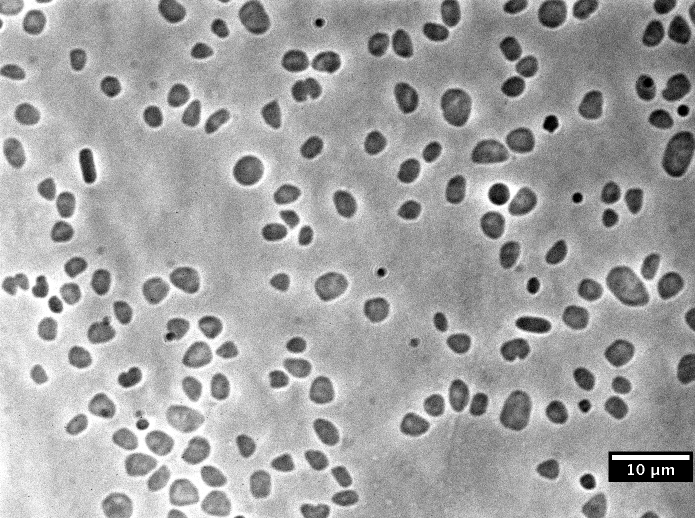
Фото взято с Wikimedia, автор - Yejineun
| Позиция | Значение |
|---|---|
| Раздел UniProtKB | Swiss-Prot |
| UniProt ID | CETZ2_HALVD |
| UniProt AC | D4GTC1 |
| EMBL AC нуклеотидной записи | CP001956; AOHU01000097 |
| PDB ID | 4B45 |
| Длина | 360 AA |
| Молекулярная масса | 37109 Дa |
| Рекомендуемое UniProt название | Tubulin-like protein CetZ2 |
| Фрагменты с известной структурой | Chain A = 1-349AA, chain B = 350-360AA |
Как видно из истории изменений записи, в феврале-марте 2015 года она была перенесена из TrEMBL в Swiss-Prot, то есть вручную проверена. Семейство CetZ не представлено в эукариотических организмах, зато в бактериях членов этого семейства много. В семейство CetZ также входит множество ГТФаз, участвующих в делении клеток. Помимо белков данного семейства в контроль формы клеток архей вовлечены кренактин и MreB. Интересно заметить, что всего 3 белка из семейства CetZ были зарегистрированы в экспериментах.
Кластерный анализ белка CetZ2 из Haloferax volcanii показал его малую распространённость среди других видов живых организмов. Так, в кластер UniRef100_D4GTC1 входит только сам изучаемый белок и полностью содержащаяся в нём последовательность из UniParc, относящаяся к той же архее. Кластер UniRef90_D4GTC1 содержит 32 белка из разных архей рода Haloferax, сгруппированных по двум семействам: собственно CetZ и белки, принимающие участие в делении клетки. Кластер UniRef50_D4GTC1 содержит 372 белка из различных видов архей, сгруппированные по тем же двум семействам. Во всех кластерах изучаемый белок является репрезентативным, а в UniRef100 и UniRef90 - ещё и сидом.
| Организм | Haloferax volcanii | Methanosarcina acetivorans |
|---|---|---|
| Cluster ID | UP000008243 | UP000002487 |
| Число белков, в том числе: | 3911 | 4468 |
| из Swiss-Prot | 183 | 519 |
| трансмембранные | 288 | 937 |
| ферменты | 964 | 517 |
| вовлечены в контроль формы клетки | 6 | 1 |
Как видно, в архее Haloferax volcanii намного шире, чем в Methanosarcina acetivorans, представлены белки, вовлечённые в контроль формы клетки. Скорее всего, это связано с тем, что изучаемая архея имеет довольно сложную блюдцевидную форму [9]. Что примечательно, все обнаруженные белки принадлежат к семейству CetZ.
- Duggin I G et al, "CetZ tubulin-like proteins control archaeal cell shape", Nature, 2014
- Структура в базе данных PDB
- Страница белка на UniProt
- Тубулиновый/FtsZ-подобный ГТФазный домен в базе данных InterPro
- Альбертс, Брей, Льюис, "Молекулярная биология клетки", том 2, глава 11
- Osawa M, Anderson D, Erickson H, "Reconstitution of Contractile FtsZ Rings in Liposomes", Science, 2009
- Zaigler A, Schuster S C, Soppa J, "Construction and usage of a onefold-coverage shotgun DNA microarray to characterize the metabolism of the archaeon Haloferax volcanii", Molecular Microbiology, 2003
- Albers S-V, Meyer B H, "The archaeal cell envelope", Nature Reviews Microbiology, 2011
- Гэррити, Буне, Кастенхольц, "Справочник Берджи по бактериологической систематике", том 1