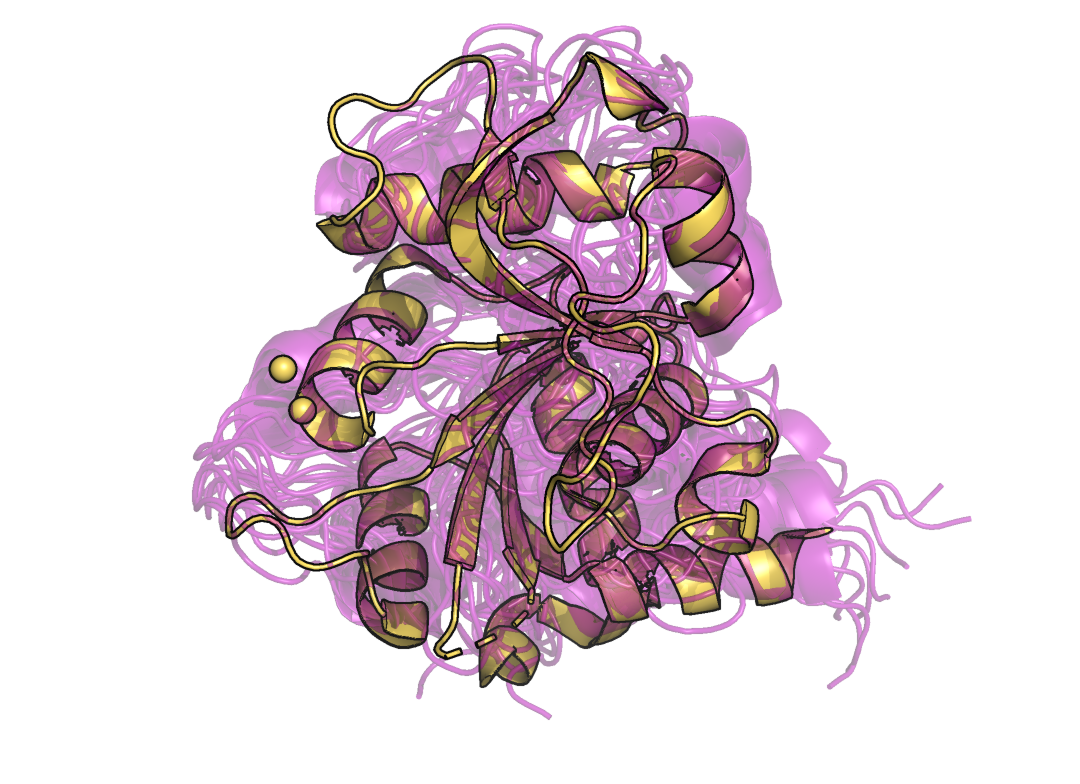
Были даны структуры были даны две структуры одного и того же белка (3,4-ДИГИДРОКСИ-2-БУТАНОН 4-ФОСФАТ СИНТАЗА): 1IEZ - структура, полученная с помощью ЯМР (10 моделей) и 1G57, полученную с помощью PCA (разрешение 1,4 Å). Сначала сопоставим эти структуры(Рис.1)(фиолетовый - структура 1IEZ, желтый - структура 1G57).
Рисунок 1. Фиолетовый - структура 1IEZ(ЯМР), желтый - структура 1G57(РСА).
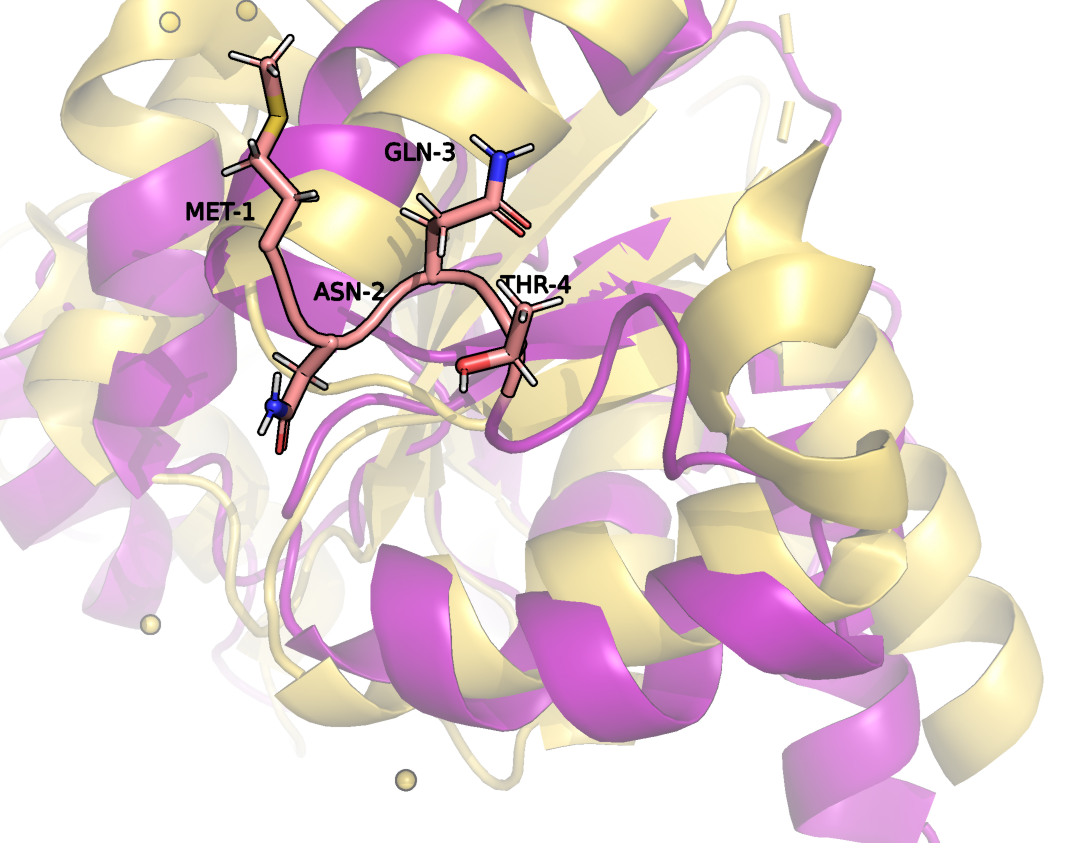
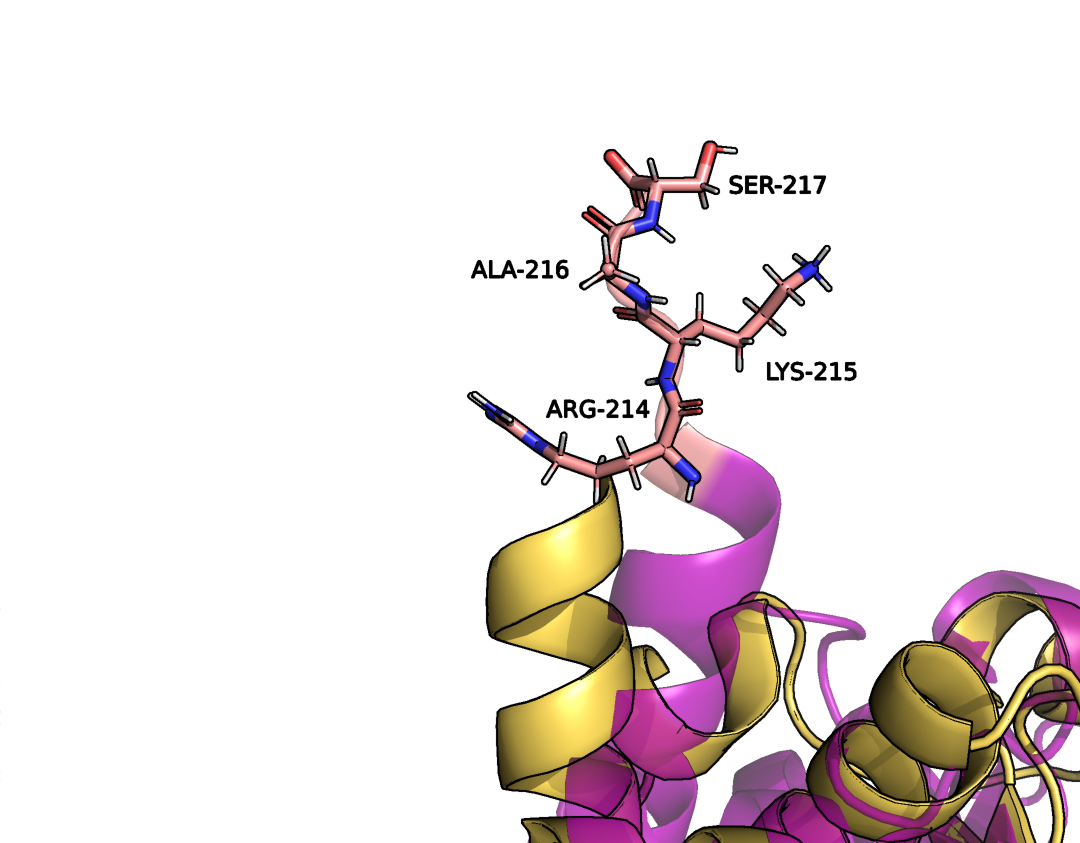
Как можно заметить, структуры неплохо сопоставляются друг другу, однако в моделях, полученных методом ЯМР нет молекул растворителя, в отличии от РСА модели. Это связано с тем, что в ЯМР используют дейтериновые растворители. Кроме того, различия заметны в наиболее подвижных участках белка: в РСА модели отсутствуют 4 первых и 4 последних аминокислотных остатка, а так же остатки под номерами 34-37 (Рис. 2-4)(отсутствующие в РСА модели остатки выделены розовым).
Рисунок 2. Начальный участок цепи.
Рисунок 3. Остатки под номерами 34-37.
Рисунок 4. Концевой участок цепи.
Вероятнее всего, эти остатки не имеют строгого положения и методом РСА их определить не удается, поскольку в разных участках цепи они занимают разное положение в разных ячейках.
Кроме того, в структуре РСА присутствует 2 цепи, между которыми можно посмотреть взаимодействия, а также молекулы лиганда - цезия. Также хочется отметить, что в структуре, полученной посредством РСА нет водородов (разрешение недостаточно хорошее).
Посмотрим некоторые взаимодействия в структуре 1G57:
1. Взаимодействие с лигандом Cs+ (фиолетовый) (Рис. 5)
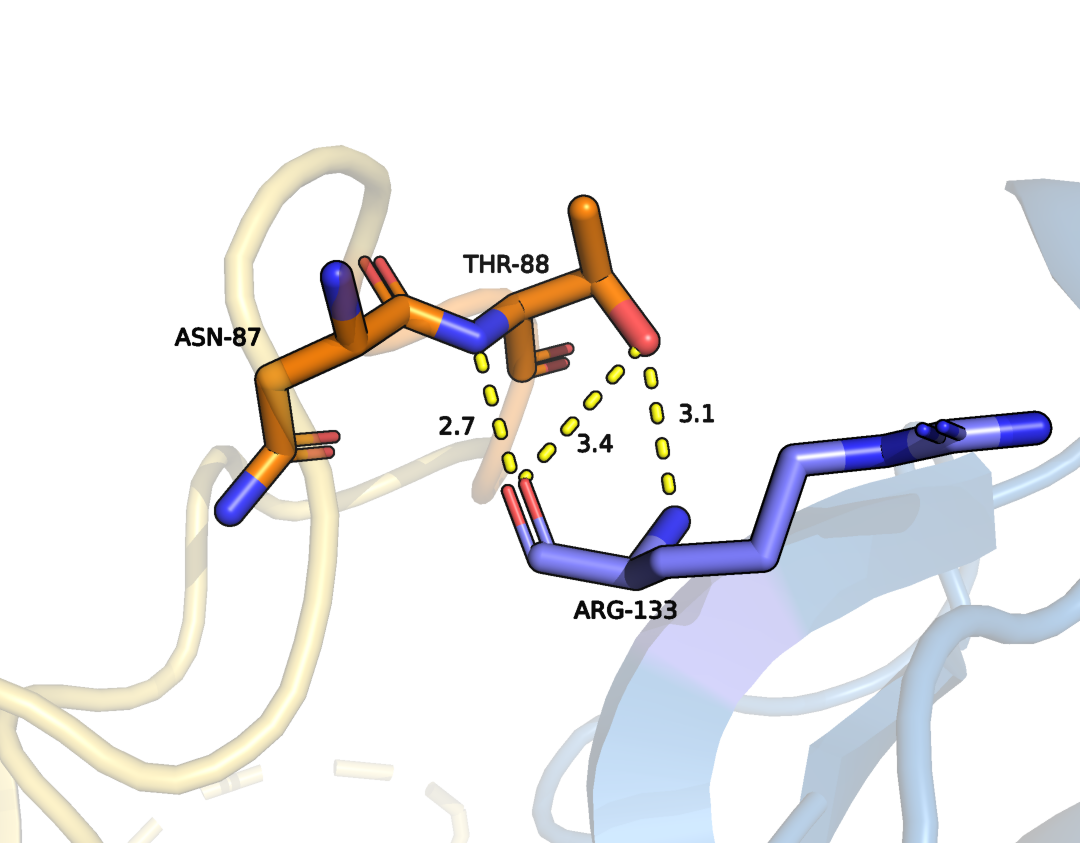
2. Водородные связи между THR88, ASN87 цепи А и ARG133 цепи В (Рис. 6)
Рисунок 5. Взаимодействия с лигандом 1G57. Цепь А отмечена оранжевым, связи с ней - зеленым, цепь В - синий, связи - желтый.
Рисунок 6. Водородные связи между THR88, ASN87 цепи А и ARG133 цепи В.
RMSF
Запись в PDB, полученная методом ЯМР, представляет собой набор моделей, которые, по мнению авторов, являются лучшими "догадками" о реальной структуре. Это происходит из-за неполноты данных об ограничениях на расстояния и ковалентную связность, получаемых из эксперимента.
Предположим, что наши модели действительно отражает вариацию в позициях, которая появляется вследствие подвижности белка во времени. Мерой подвижности отдельных его участков (атомов, остатков, петель) является RMSF. В идеальном случае, если наш ансамбль в действительности представляет собой отражение эволюции системы во времени, по значениям RMSF атомов даже вычисляют значения B-факторов аналитически. Таким образом, у нас есть возможность грубо прикинуть, в какой мере ансамбль моделей в записи PDB, полученной методом ЯМР, действительно можно принять за отражение подвижности белка, взяв для сравнения значения B-факторов из модели РСА.
В этом задании было интересно построить график зависимости величины B-фактора для структуры РСА от величины RMSF для структур ЯМР того же белка (Рис. 7).
Рисунок 7. Зависимость RMSFs, полученных по данным структуры ЯМР от средних значений В-факторов, полученных по РСА структуре.
Значения RMSF рассматривались только для тех остатков, которые есть в структуре ЯМР (5-33;38-213). Как мы видим, значения RMSF с В-фактором соотносятся: остатку с большим B-фактором соответствует большее значение RMSF, однако говорить о характере зависимости сложно, хотя она явно присутствует (если посмотреть корреляции, то получается: по Пирсону - rho=0.54, p-val=1.27e-16 и по Спирману - rho=0.46, p-val=6.38e-12).
ЯМР vs РСА на уровне отдельных связей
Для каждой из предложенных категорий водородных связей были выбраны следующие (в РСА-структуре):
1. Водородная связь между атомами остова в ядре белка (в альфа-спирали или бета-листе):
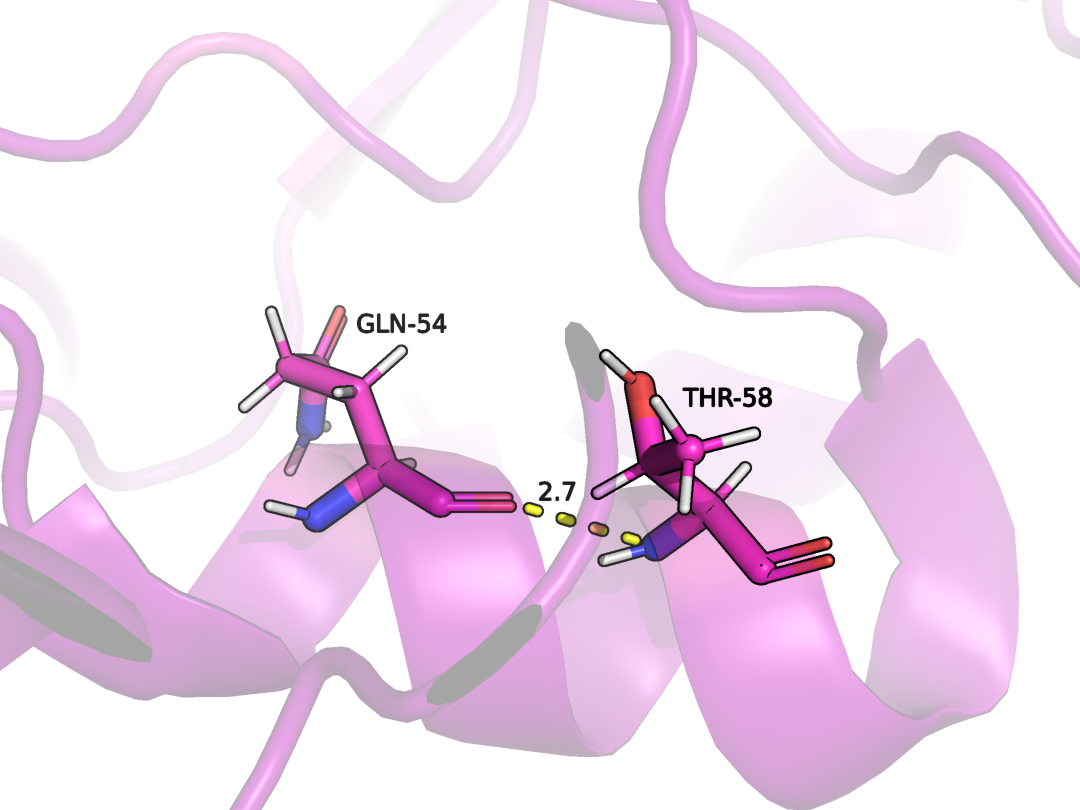
GLN54 и THR58 в цепи А
2. Водородная связь боковых цепей в ядре белка:
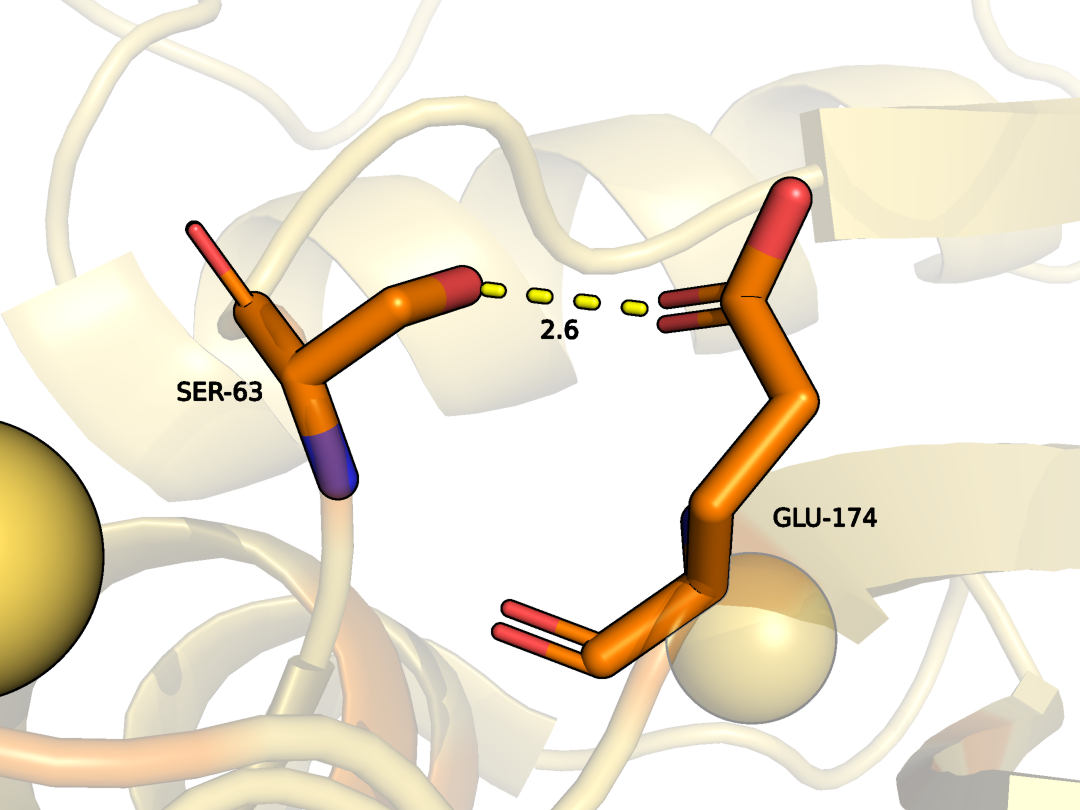
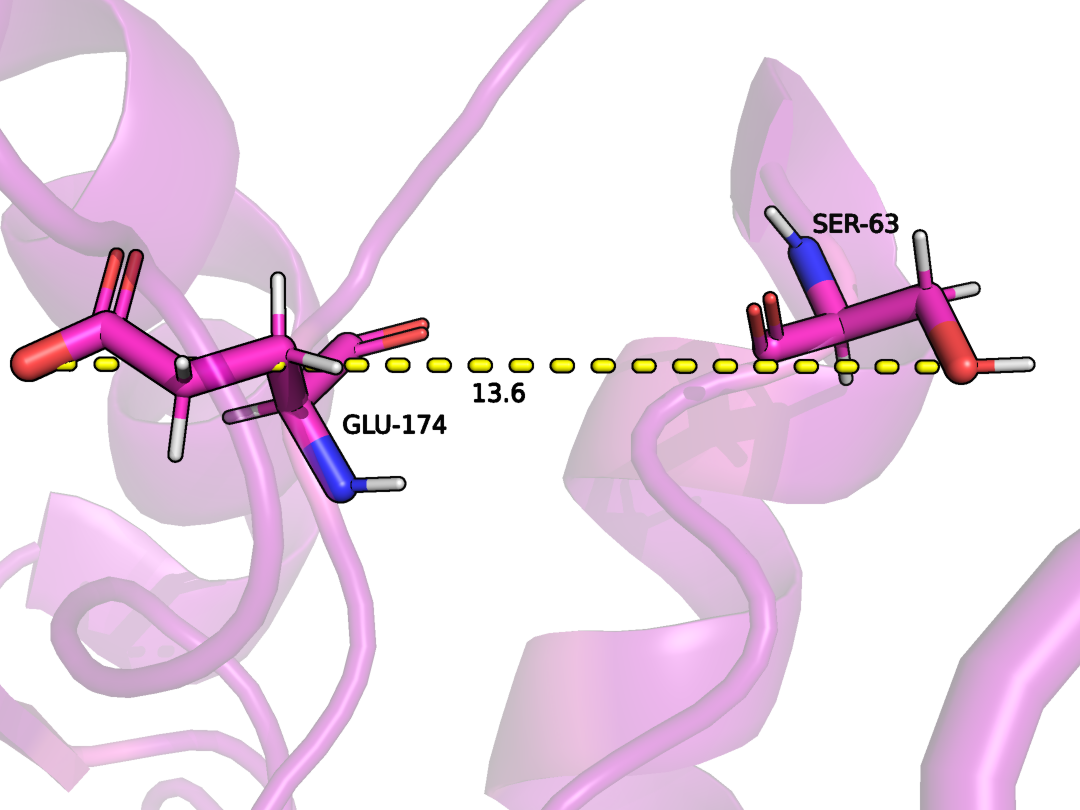
SER63 и GLU174 цепи А (были взяты ближе к периферии)
3. Водородная связь в петлях, выходящих на поверхность глобулы:
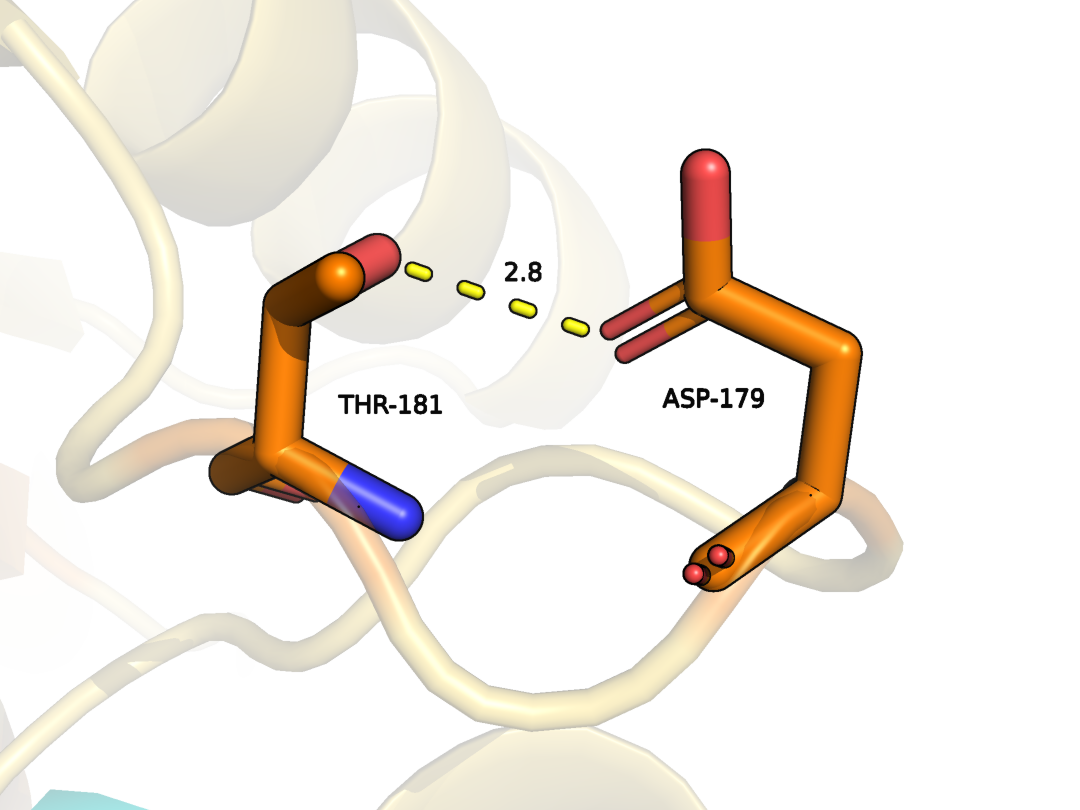
THR181 и ASP179 цепи А
Водородная связь между атомами остова в ядре белка
Данные остатки находятся в альфа-спирали, поэтому не удивительно, что эта связь присутствует во всех ЯМР моделях. Данная спираль не сильно подвергается изменениям, поэтому общая конформация меняется не сильно (Рис. 8-9)
Рисунок 8. Водородная связь между GLN54 и THR58 в цепи А в модели РСА.
Рисунок 9. Водородная связь между GLN54 и THR58 в цепи А в модели ЯМР.
Водородная связь боковых цепей в ядре белка
Можно заметить, что в структуре ЯМР эти остатки ориентированы в разны стороны, поэтому взаимодействий между ними не наблюдается. (Рис. 10-11). Если смотреть на значения параметров Ramachandran outliers и Sidechain outliers для структуры РСА и ЯМР, то для РСА значения будут 0% (несколько сомнительно, но допустим) и 3,4% соответственно, что сильно меньше значений для структуры ЯМР - 7.1% и 29.9% соответственно. Поэтому в данном случае я больше склонна доверять структуре РСА в целом.
Рисунок 10. Водородная связь между SER63 и GLU174 цепи А (были взяты ближе к периферии) в модели РСА.
Рисунок 11. Водородная связь между SER63 и GLU174 цепи А (были взяты ближе к периферии) в модели ЯМР.
Водородная связь в петлях, выходящих на поверхность глобулы
В структурах ЯМР эти остаткри в большинстве случаев повернуты таким образом, что связи между ними не наблюдается. (Рис. 12-13)
Рисунок 12. Водородная связь между THR181 и ASP179 цепи А в модели РСА.
Рисунок 13. Водородная связь между THR181 и ASP179 цепи А в модели ЯМР.
В целом можно сказать, что связи в подвижных структурах белка воспроизводятся в ЯМР довольно плохо, вторичные структуры, наоборот, строго организованные части белка, поэтому и водородные связи в ЯМР структурах в большинстве случаев совпадают с таковыми в РСА структурах.
Ниже приведена сводная таблица (Таблица 1).
Таблица 1. Примеры водородных связей в структуре белка .
Положение в структуре
Связанные остатки
Расстояние в РСА, Å
% моделей ЯМР со связью
Минимальное расстояние в ЯМР, Å
Максимальное расстояние в ЯМР, Å
Медиальное расстояние в ЯМР Å
Mежду атомами остова в ядре белка
GLN54 и THR58 в цепи А
3.0
100
2.7
3.6
3.35
Между атомами боковых цепей в ядре белка
SER63 и GLU174 цепи А
2.6
0
6.8
14.4
10.6
Между атомами в петлях, выходящих на поверхность глобулы