На главную страницу третьего семестра.
Занятие 6. Работа с Chemketch.
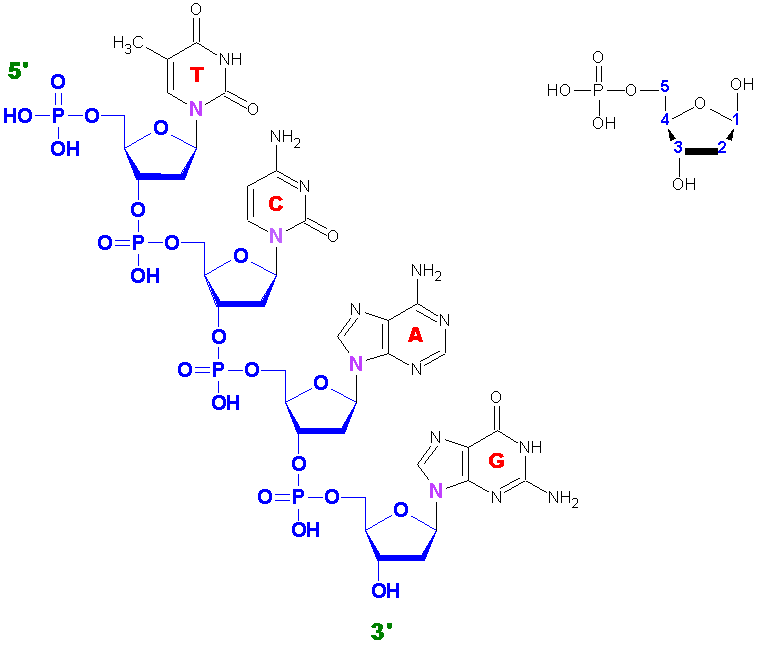
Задание 1. Построение цепи ДНК.
С помощью программы СhemSketch была построена цепь ДНК, состоящая из 4-х нуклеотидов T,C,A,G. Атомы дезоксирибозы перенумированы на отдельном примере сахара, дабы не загромождать общий вид цепочки. Выделены синим цветом те химические связи, которые описывают ход сахаро-фосфатного остова. Выделены сиреневым азоты оснований, связанные с остовом. Подписаны 3'- и 5'-концы цепи.
Задание 2. Химическая структура уридина и псевдоуридина.
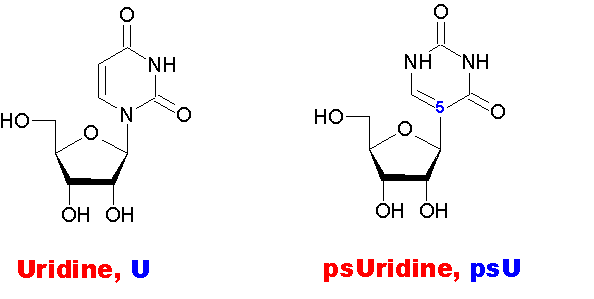
Как видно на рисунке, псевдоуридин отличается от уридина только тем, что гликозидная связь образуется не через атом N(1), как в уридине, а через атом С(5). Таким образом, данные рибонуклеозиды - структурные изомеры.
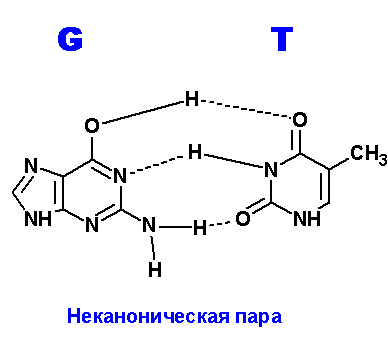
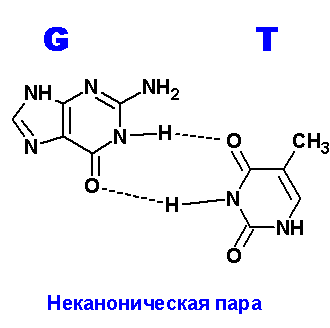
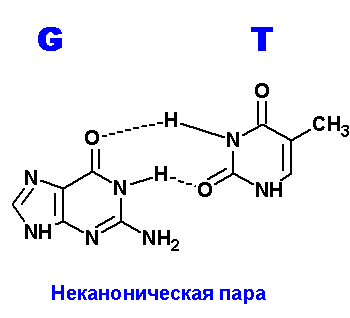
Задание 3. Водородные связи между пуриновыми (A,G) и пиримидиновыми (T,C) основаниями.
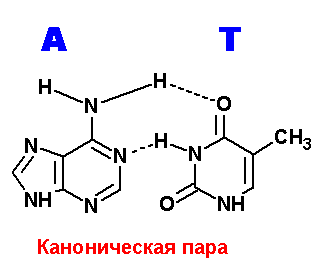
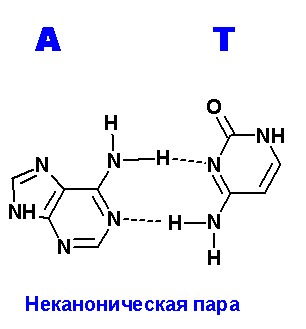
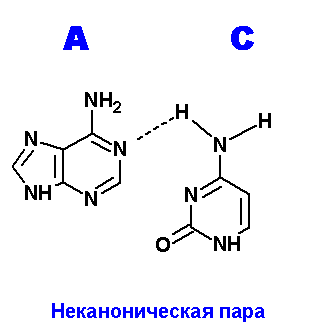
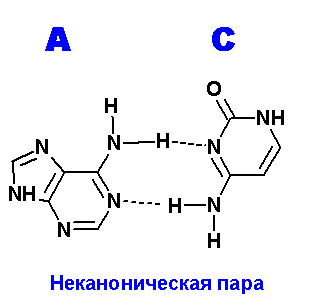
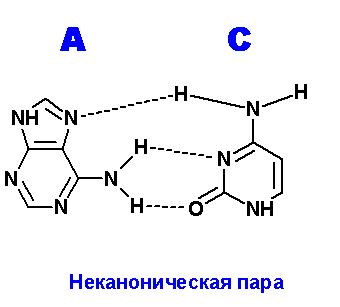
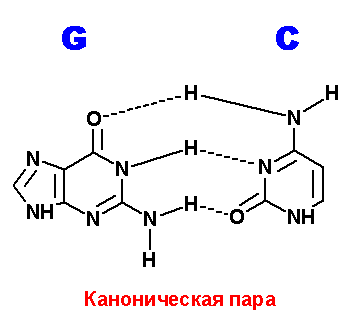
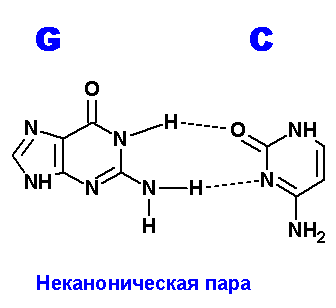
Ниже приведены все теоретически возможные и (невозможные) структуры пар нуклеотидов (только пурины с пиримидинами - всего четыре варианта взаимодействий). Так как всего таких пар можно придумать довольно много, поэтому для изображения использовались только те пары, контакты между которыми осуществляются по-крайней двумя водородными связями. Итак, вот эти структуры:
Очевидно, наиболее крепкие и вероятные пары A=T.
Очевидно, наиболее крепкие и вероятные пары A=С. Понятно, что все эти пары будут неканоническими и довольно редко встречаемыми в природе (если вообще такое существует) из-за сильных конформационных изменений, претерпеваемых молекулой ДНК, например, в сахаро-фосфатном остове какие-нибудь изломы, при попытке включить данные пары в спираль.
Наиболее крепкие и вероятные пары G=С. В данном случае, сложно предположить что-то лучшее, чем каноническая пара, или хотя бы близкое к этой паре. Но все же можно нарисовать одну пару с двумя водородными связями. Правда, атомы азота, образующие гликозидую связь с остатком пентозы, оказываются по разные стороны от области контакта. А это очень не выгодно с точки зрения конформационной структуры ДНК.
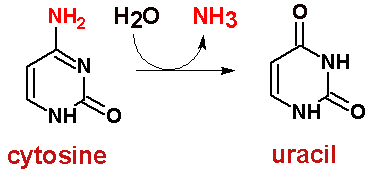
Наиболее крепкие и вероятные пары G=T. Число неуотсон-криковских пар можно предложить побольше и количество водородных связей в этих парах также велико. Вполне возможно, что это явление (довольно крепкие контакты) является причиной многих точечных мутаций в ходе репликации генома. Действительно, по количеству донорных и акцепторных атомов, тимин ничем не отличается от урацила. То есть, все структуры, нарисованные для тимина в контакте с гуанином, можно с тем же успехом применить к парам урацил:гуанин (U:G). Но, как известно, экспериментально доказано, что цитозин, также как и пурины, может спонтанно дезаминироваться. Но тогда получится не что иное, как УРАЦИЛ!!
То есть, появляется гибридная пара G-U, которая, если не будет исправлена, может привести к мутации (после репликации в одной цепочке ДНК также, как и в материнской ДНК, пара G:C, тогда как в другой цепочке ДНК - возникнет пара T:A (привожу пример, когда уже после репликации все цепочки прометилированы, как известно для того, чтобы избегать многих досадных ошибок в ходе репликации и также для выявления встроившегося вирусного генома, если такое произошло, системой рестрикции/репариции хозяина)).
©Володя Рудько