Вторичная структура РНК
Предсказание вторичной структуры РНК осуществлялось двумя методами: с помощью поиска инвертированных повторов (einverted из emboss) и с помощью алгоритма Зукера, который реализуется с помощью программы RNAfold из пакета Viena RNA Package.
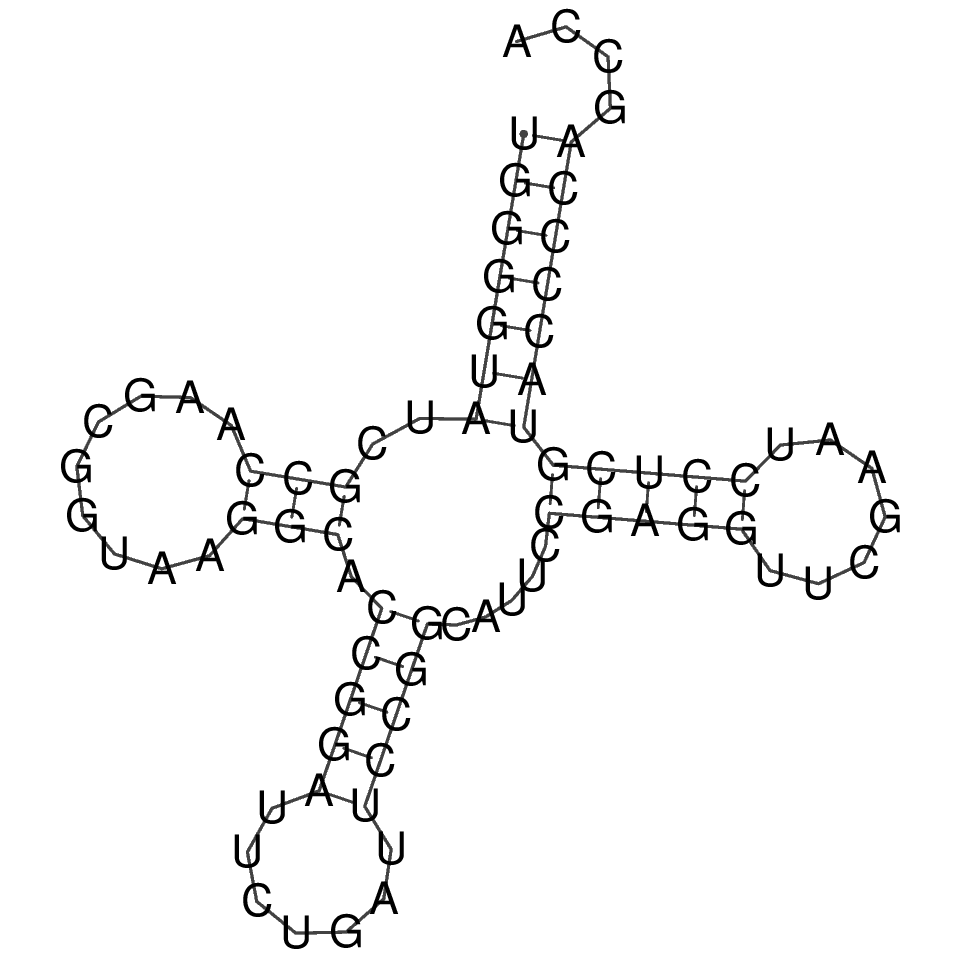
Изображение вторичной структуры РНК, полученное с помощью пакета Viena RNA Package
| Участок структуры | Позиции в структуре (find_pair) | Предсказание с помощью einverted | Предсказание по алгоритму Зукера |
| Акцепторный стебель | 5'_2-7_3'; 5'_66-71_3'; 6 пар | 6/6 | 6/6 |
| D-стебель | 5'_10-12_3'; 5'_23-25_3'; 3 пары | 0/3 | 3/3 |
| T-стебель | 5'_49-53_3'; 5'_61-65_3'; 5 пар | 0/5 | 5/5 |
| Антикодоновый стебель | 5'_27-31_3'; 5'_39-43_3'; 5 пар | 4/5 | 5/5 |
| Общее число канонических пар нуклеотидов | 19 | 10/19 | 19/19 |
Как видно из данных в таблице, алгоритм Зукера намного лучше подходит для предсказания вторичной структуры РНК.
ДНК-белковые контакты
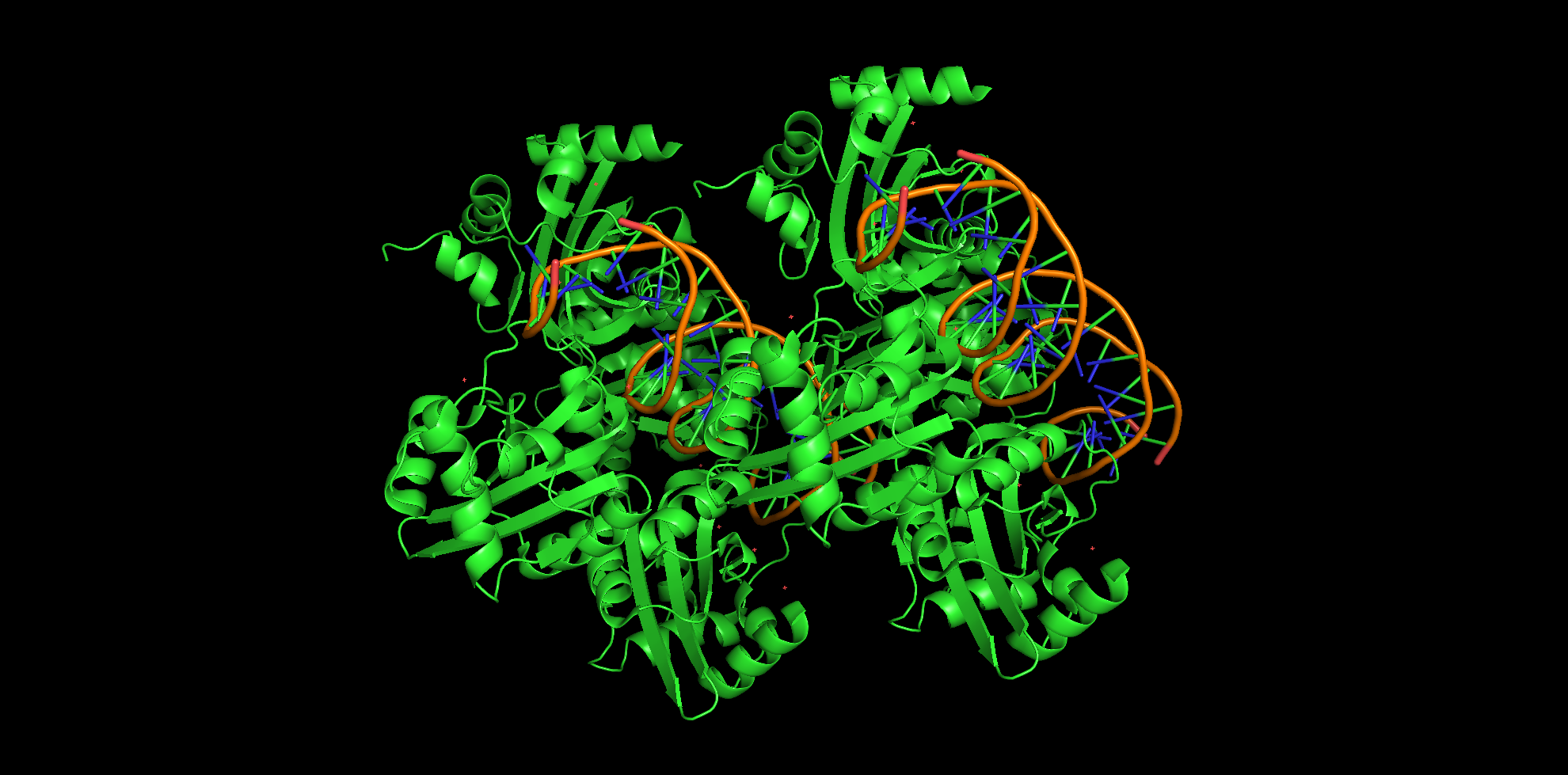
В процессе выполнения данного задания для ДНК-белкового комплеса из вируса папилломы были проанализированы полярные и неполярные контакты между аминокислотными остатками и нуклеиновыми кислотами. Анализ производится в программе PyMol и программе Nucplot.
 |
Описание контактов | |||
| Место контакта с белком | Полярные контакты | Неполярные контакты | Общее число контактов | |
| 2'-дезоксирибоза | 0 | 23 | 23 | |
| Фосфорная кислота | 4 | 10 | 14 | |
| Малая бороздка ДНК | 0 | 8 | 8 | |
| Большая бороздка ДНК | 0 | 4 | 4 | |
Число контактов было посчитано для 2 белковых цепей, так как одной молекулой ДНК с одной стороны взаимодействуют 2 молекулы, а не одна.
Общее число контактов между белком и ДНК очень незначительно. Большая часть из них (~92%) приходится на неполярные контакты, причем большая половина из них осуществляется через остаток дезоксирибозы. Полярные контакты в небольшом количестве нашлись между остатками форфорной кислоты и аминокислотами (всего 4 штуки). Контакты с малой бороздкой ДНК встречаются в 4 раза чаще, чем с большой.
Из этого анализа можно сделать вывод, что в данной структуре нет очень прочной связи между ДНК и белком. Также можно судить о неспецифичности ДНК-белкового контакта, что будет проиллюстрировано ниже.
Скачать скрипт для PyMol с определениями множеств
Скачать скрипт для PyMol с выделениями по множествам
Скачать скрипт для PyMol, использовавшийся для подсчета контактов
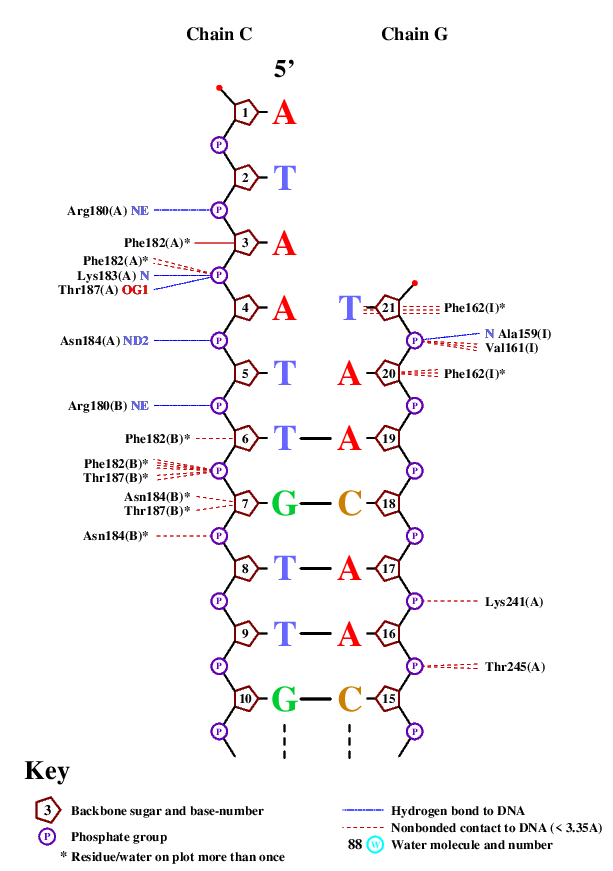
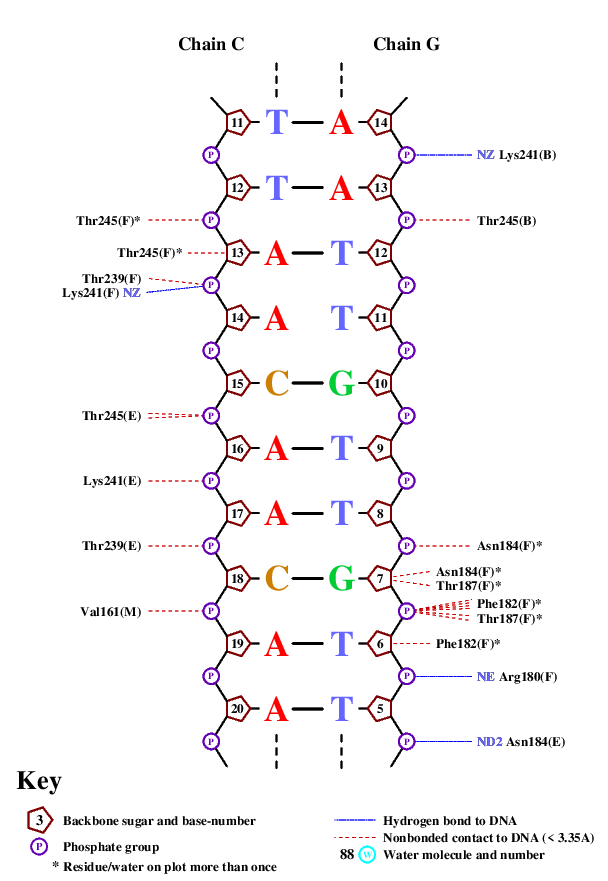
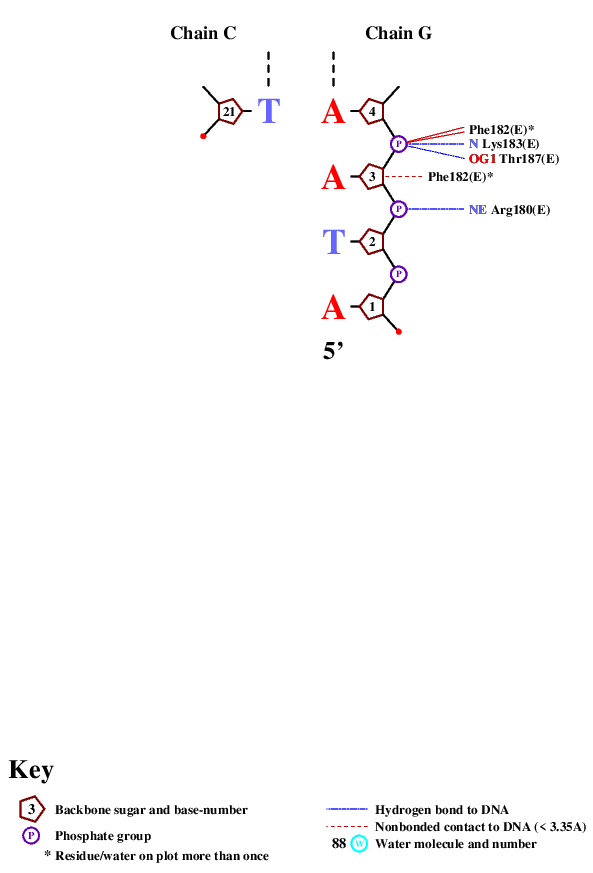
Выше приведен список контактов, которые находит программа nucplot. Как видно на изображениях, основным местом межмолекулярного контакта являются остатки фосфорной кислоты и 2'-дезоксирибозы. Непосредственно с азотистым основанием взаимодействует единственная аминокислота. Этот факт подтверждает предположение, что данное взаимодействие не является специфичным по отношению к ДНК.
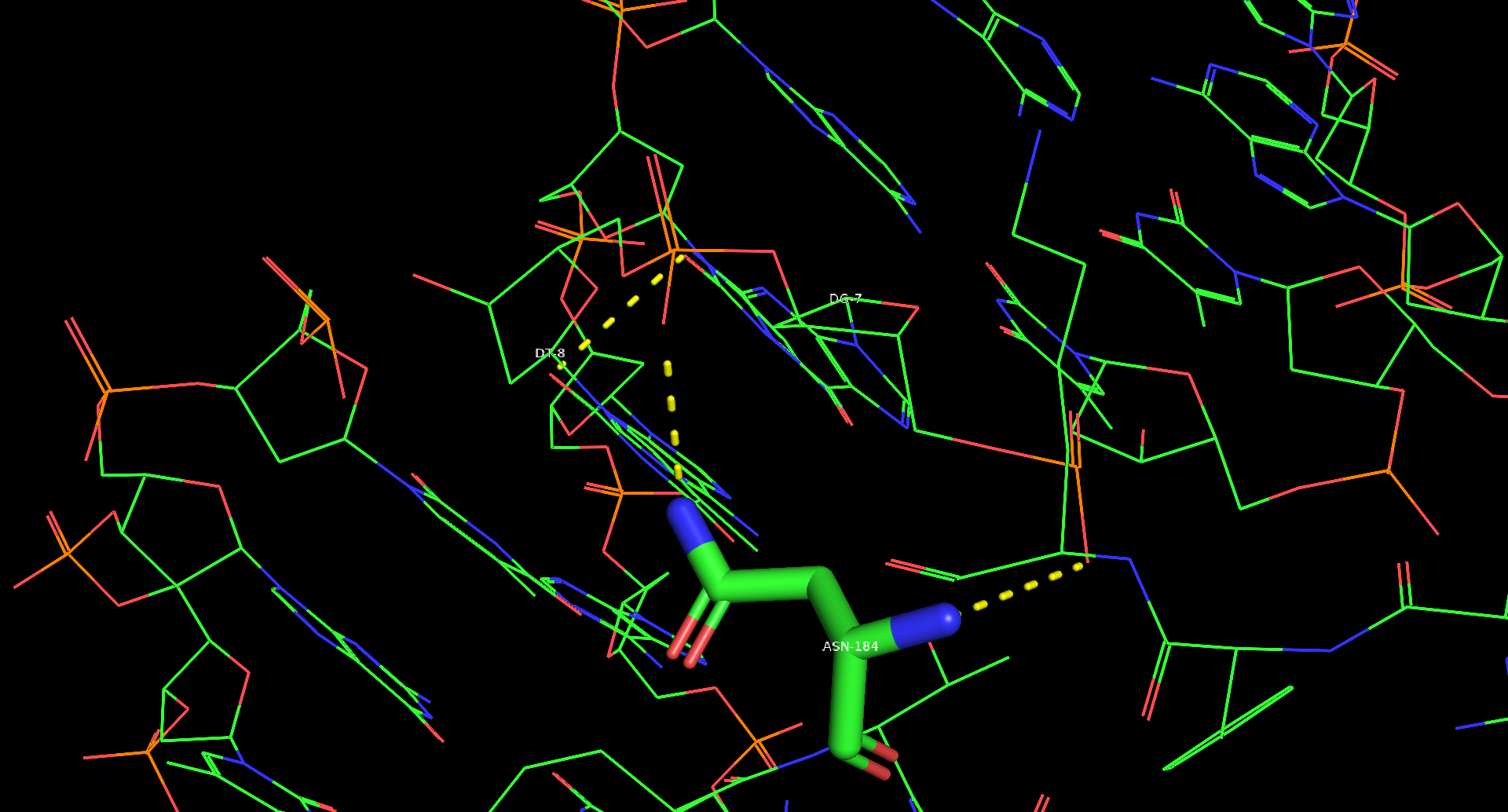
Наибольшее количество контактов с ДНК имеет 184 остаток аспарагина на каждой из белковых цепей. Были проанализированы контакты одного такого остатка. С помощью функции PyMol "find polar contacts" были найдены 2 полярных контакта для этого остатка. Аспарагин взаимодействует с кислородом остатка фосфорной кислоты.
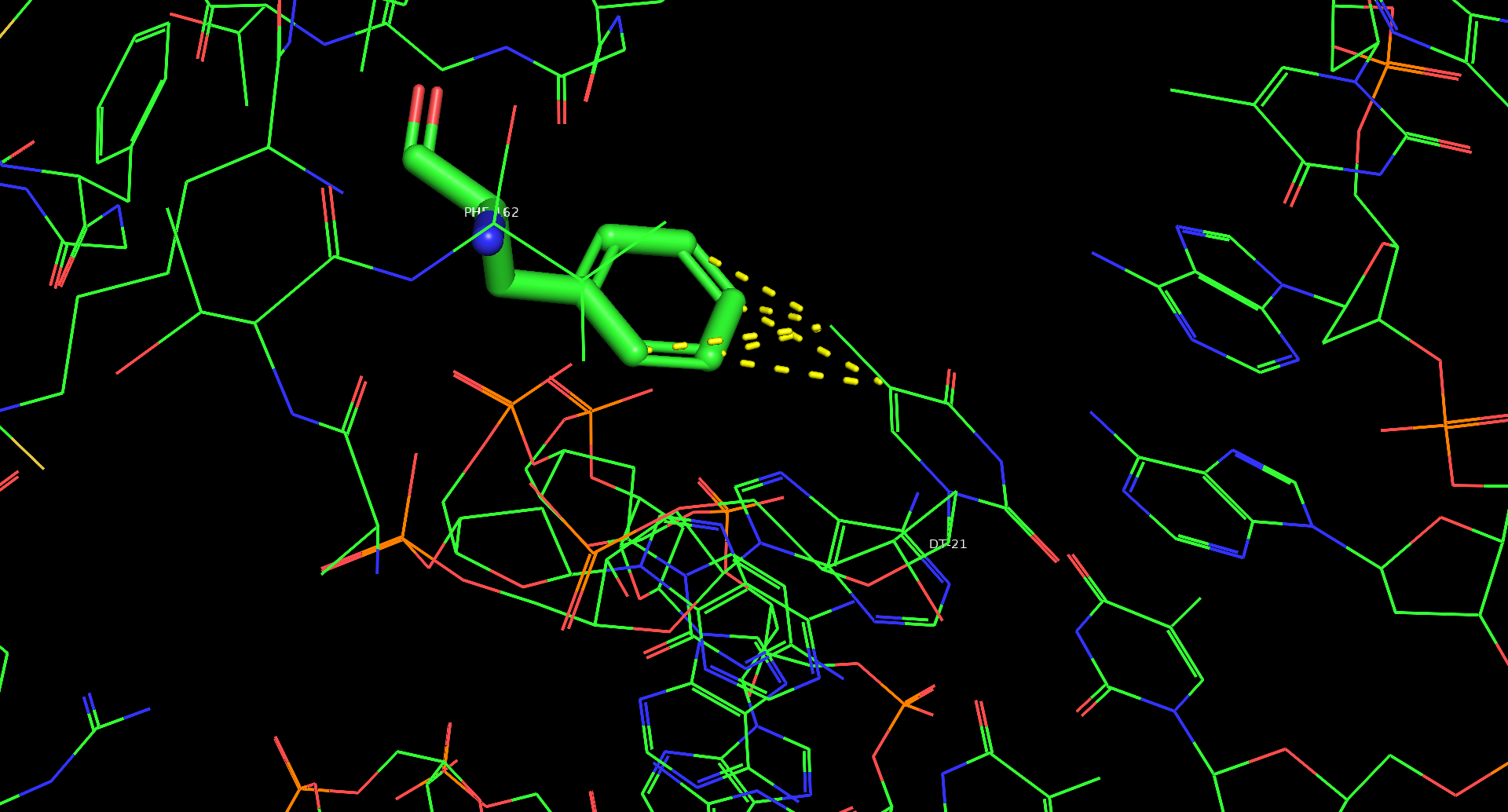
Для остатка Phe162(I) показаны контакты с азотистым основанием ДНК и именно этот остаток может принимать участие в узнавании ДНК. Остаток контактирует с тимином, причем растояние между аминокислотой и основанием не превышает 3,5Å, что позволяет предположить стекинг-взаимодействия между данными структурами. Исходя из данных, полученных с помощью программы nucplot, этот же остаток фенилаланина взаимодействует с дезоксирибозой следующего нуклеотида, что подчеркивает важность этого межмолекулярного контакта.