| Название
| Название (en)
| Однобуквенное обозначение
| Трехбуквенное обозначение
| Формула
| Распространенность
| Неводородных атомов в боковой цепи
| Размер
| Гидрофобный
| Гидрофильный
| Особенности
| Ароматичность
| Заряд
| Донор вод. связей
| Акцептор вод. связей
| Донор вод. cвязей (боковой цепью)
| Акцептор вод. cвязей (боковой цепью)
| Образует дисульфидные связи
| Стерические ограничения
|
| Аланин |
Alanine |
A |
ALA |
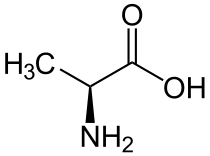 |
часто |
1 |
маленький |
да |
нет |
|
нет |
0 |
да |
да |
нет |
нет |
нет |
нет |
| Цистеин |
Cysteine |
C |
CYS |
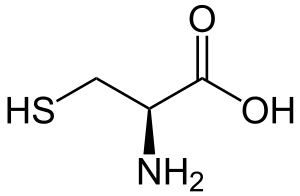 |
редко |
2 |
маленький |
да |
нет |
боковая цепь может быть донором водородных связей; образует дисульфидные связи |
нет |
0 |
да |
да |
да |
нет |
да |
нет |
| Аспартат (аспарагиновая кислота) |
Aspartate (Aspartic acid) |
D |
ASP |
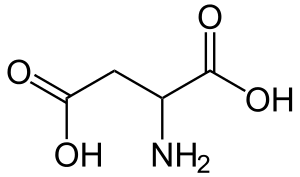 |
часто |
4 |
средний |
нет |
да |
отрицательно заряженный; боковая цепь может быть акцептором водородных связей |
нет |
– |
да |
да |
нет* |
да |
нет |
нет |
| Глутамат (Глутаминовая кислота) |
Glutamate (Glutamic acid) |
E |
GLU |
 |
часто |
5 |
средний |
нет |
да |
отрицательно заряженный; боковая цепь может быть акцептором водородных связей |
нет |
– |
да |
да |
нет* |
да |
нет |
нет |
| Фенилаланин |
Phenylalanine |
F |
PHE |
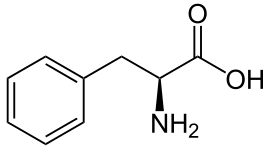 |
редко |
7 |
большой |
да |
нет |
ароматический |
да |
0 |
да |
да |
нет |
нет |
нет |
нет |
| Глицин |
Glycine |
G |
GLY |
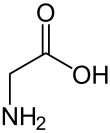 |
часто |
0 |
маленький |
нет |
да |
более слабые, чем для других а.к. ограничения на торсионные углы остова, часто встречается в поворотах полипептидной це |
нет |
0 |
да |
да |
нет |
нет |
нет |
более слабые, чем для других а.к. ограничения на торсионные углы остова, часто встречается в поворотах полипептидной цепи |
| Гистидин |
Histidine |
H |
HIS |
 |
средне |
6 |
большой |
нет |
да |
ароматический; положительно заряженный; боковая цепь может быть донором водородных связей |
да |
+ |
да |
да |
да |
нет* |
нет |
нет |
| Изолейцин |
Isoleucine |
I |
ILE |
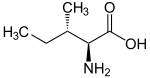 |
часто |
4 |
средний |
да |
нет |
|
нет |
0 |
да |
да |
нет |
нет |
нет |
нет |
| Лизин |
Lysine |
K |
LYS |
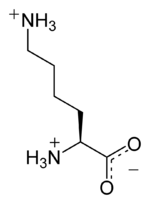 |
средне |
5 |
средний |
да |
да |
положительно заряженный; боковая цепь может быть донором водородных связей |
нет |
+ |
да |
да |
да |
нет* |
нет |
нет |
| Лейцин |
Leucine |
L |
LEU |
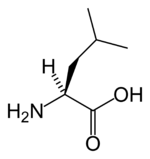 |
часто |
4 |
средний |
да |
нет |
|
нет |
0 |
да |
да |
нет |
нет |
нет |
нет |
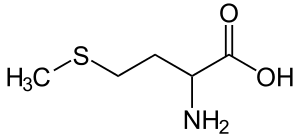
| Метионин |
Methionine |
M |
MET |
 |
редко |
4 |
средний |
да |
нет |
|
нет |
0 |
да |
да |
нет |
нет |
нет |
нет |
| Аспарагин |
Asparagine |
N |
ASN |
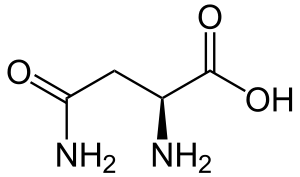 |
часто |
4 |
средний |
нет |
да |
боковая цепь может быть донором водородных связей; боковая цепь может быть акцептором водородных связей |
нет |
0 |
да |
да |
да |
да |
нет |
нет |
| Пролин |
Proline |
P |
PRO |
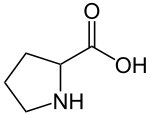 |
редко |
3 |
маленький |
да |
нет |
не может быть донором водородных связей; жестко зафиксированные торсионные углы остова, характерные для α-спиралей; часто встречается в первом витке спира |
нет |
0 |
только на N-конце (и поэтому нехарактерен для центральной части спиралей и тяжей) |
да |
нет |
нет |
нет |
жестко зафиксированные торсионные углы остова, характерные для α-спиралей; часто встречается в первом витке спирали |
| Глутамин |
Glutamine |
Q |
GLN |
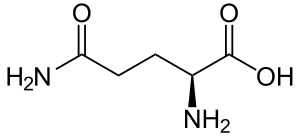 |
часто |
5 |
средний |
нет |
да |
боковая цепь может быть донором водородных связей; боковая цепь может быть акцептором водородных связей |
нет |
0 |
да |
да |
да |
да |
нет |
нет |
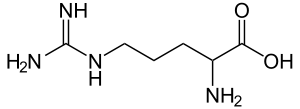
| Аргинин |
Arginine |
R |
ARG |
 |
средне |
7 |
большой |
да |
да |
положительно заряженный; боковая цепь может быть донором водородных связей |
нет |
+ |
да |
да |
да |
нет* |
нет |
нет |
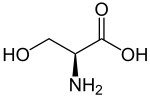
| Серин |
Serine |
S |
SER |
 |
часто |
2 |
маленький |
нет |
да |
боковая цепь может быть донором водородных связей; боковая цепь может быть акцептором водородных связей |
нет |
0 |
да |
да |
да |
да |
нет |
нет |
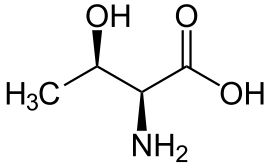
| Треонин |
Threonine |
T |
THR |
 |
часто |
3 |
маленький |
нет |
да |
боковая цепь может быть донором водородных связей; боковая цепь может быть акцептором водородных связей |
нет |
0 |
да |
да |
да |
да |
нет |
нет |
| Валин |
Valine |
V |
VAL |
 |
часто |
3 |
маленький |
да |
нет |
|
нет |
0 |
да |
да |
нет |
нет |
нет |
нет |
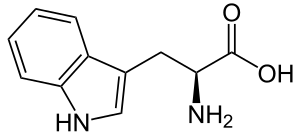
| Триптофан |
Tryptophan |
W |
TRP |
 |
очень редко |
10 |
очень большой |
да |
нет |
ароматический; боковая цепь может быть донором водородных связей |
да |
0 |
да |
да |
да |
нет |
нет |
нет |
| Тирозин |
Tyrosine |
Y |
TYR |
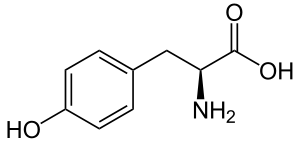 |
редко |
8 |
большой |
да |
нет |
ароматический; боковая цепь может быть донором водородных связей; боковая цепь может быть акцептором водородных связей |
да |
0 |
да |
да |
да |
да |
нет |
нет |
| |
* да в протонированной (депротонированной) форме, которая реально не встречается |