Заряд системы: -9.9999 из за фосфатных групп в остове.
Размер и формa ячейки: 5.0920 x 4.9060 x 5.49620 куб
Минимизация энергии:
- Число шагов: nsteps = 10000
Длина шага: dt = 0.001 (ps)
Алгоритм расчёта электростатики и Ван-дер-Ваальсовых взаимодействий: pme
Основной расчёт МД:
Число шагов: 5000000
Длина шага: 0.002 ps
Алгоритм интегратора: md
Алгоритм расчёта электростатики и Ван-дер-Ваальсовых взаимодействий: pme и cutt-off
Алгоритмы термостата и баростата: Berendsen
остальные параметры модели можно посмотреть тут
время первращенияв В форму t=9800.0000
Определим средне-квадратичное отколнение сначала расчитаем отклонение в ходе всей симуляции относительно стартовой структуры
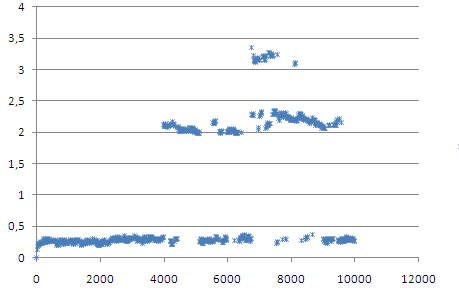
И относительно каждой предыдущей структуры на растоянии 400 кадров. Если ближе к концу закончился конформационный переход, то отколнение должно уменьшаться.
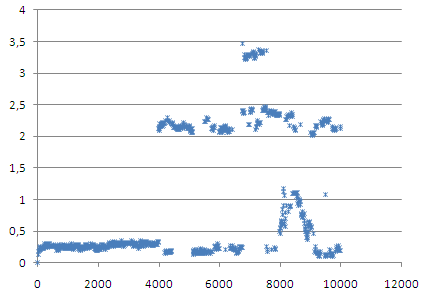
Гидрофильная и гидрофобная поверхность.
Чтобы рассмотреть, что там происходит, посмотрим только на последние 6000 шагов.
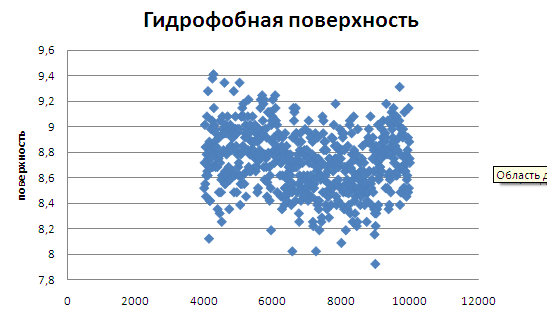
Видно, что к моменту перехода гидрофобная поверхность уменьшается.
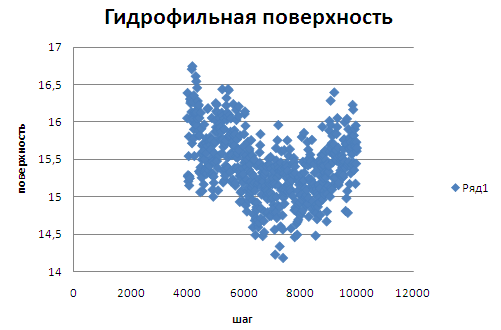
На графике гидрофильной поверхности видно более четкое уменьшение поверхности на момент перехода.
Количество водородных связей в ДНК и между ДНК и водой.
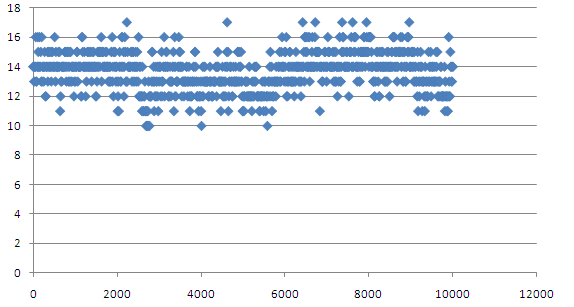
Количество водороных связей долно быть 14. Оно колеблется как раз в районе 14 в диапазоне от 10 до 16. Понятно почему этих связей может быть меньше 14, но откуда берутся лишние не ясно.
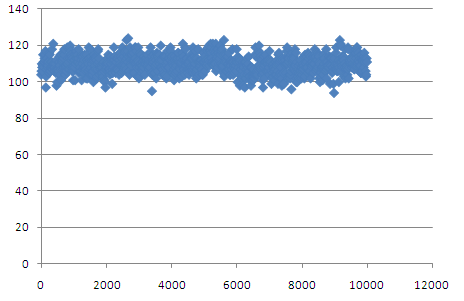
Видно, что количество воородных связей с водой колеблется от 100 до 120 и не меняется значительно в процессе перехода.
Возможно, эти связи не влият на возможность перехода из однойформы в другую. © Garanina Irina
Алогритм: integrator = l-bfgs; Goldfarb-Shanno
Алгоритм расчёта электростатики и Ван-дер-Ваальсовых взаимодействий: cut-off
Модель, которой описывался растворитель: implicit_solvent = No
Утряска растворителя: