- Краткое описание структуры в файле 1MHD.pdb
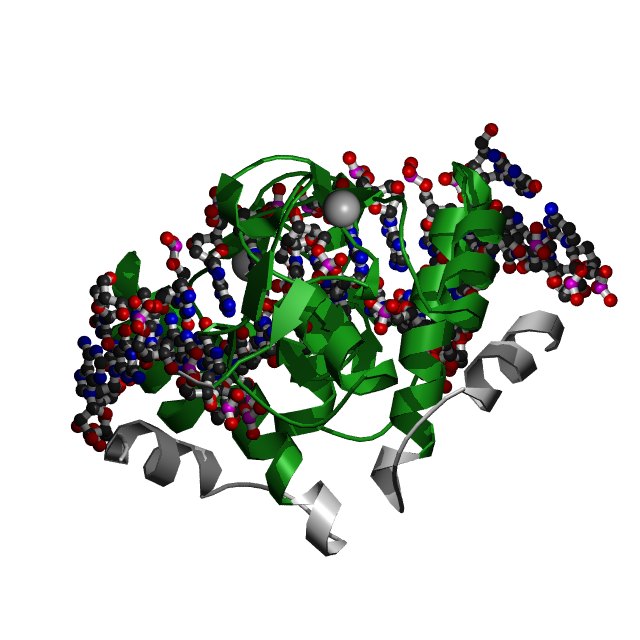
Данная структура из HOMO SAPIENS состоит из двух молекул: ДНК (цепи D и С), белок SMAD3 ( цепи А и В).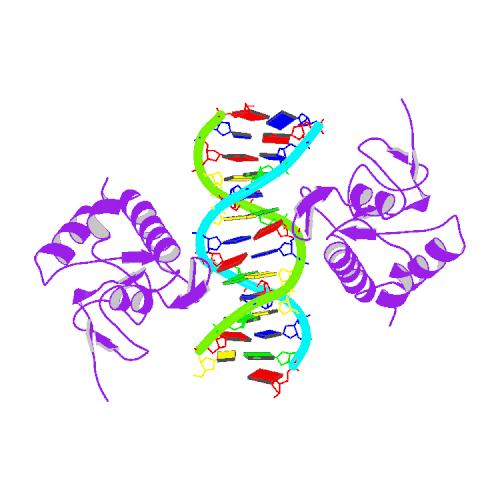
Для исследования были выбраны цепь A белка и цепи С и D, представляющие ДНК со следующей последовательностью:
цепь C [1] 5' - DC DA DG DT DC DT DA DG DA DC DA DT DA - 3' [13] | | | | | | | | | | | | | цепь D [14] 3' - DA DG DT DC DA DG DA DT DC DT DG DT DA DT - 5' [1],где 1 и 14 - номера первого и последнего нуклеотида ( согласно файлу dna_old.out цепь C нумеруется с 5` по 3`, а D - c 3` по 5`).
- Функции белка, структура которого представлена в файле 1MHD.pdb.Код структуры в Uniprot:P84022.
Цитоплазматические медиаторы-белки SMAD активируются рецептором ростовых факторов TGF-beta.
Существует несколько типов SMAD. Белок SMAD3 (MADH3) является Smad-белком,который регулируется рецептором.
Кроме того, Smad3-белки связаны с раковыми опухолевыми новообразованиями. (http://proteopedia.org/wiki/index.php/1mhd ). - Исследование структуры ДНК c помощью программ find_pair и analyze
Исходный файл программы analyze DNA_old.out
Тип данной ДНК согласно программе analyze - B.Торсионные углы A-форма B-форма Z-форма(для C) Z-форма(для G) DNA alpha -51.7 -29.9 -139.5 52.0 -52.6 beta 174.8 136.4 -136.7 179.0 179.4 gamma 41.7 31.1-31.2 50.9 -173.8 60.3 delta 79.0-79.1 143.3-143.4 137.6 94.9 141.6 epsilon(˚) -147.8 -140.8 -96.5 -103.6 -159.9 zeta -75.0- -75.1 -160.5 81.9 -64.8 -118.4 chi -157.2 -98.0 -154.3 58.7 -104.0
Cамые "деформированные" нуклеотиды ДНК согласно таблице nuclacid.xls:12Т,10С цепи 1. Для цепи 2 это нуклеотиды 6А и 7Т,однако их суммарные средние отклонения от средних значений торсионных углов значительно меньше таких же отклонений для 10С и 12Т цепи 1.
В данном случае "деформированные" нуклеотиды цепи 1 не связаны с белком, потому их деформация не объяснима связыванием с белком.Деформацию нуклеотида 6А цепи 2 можно объяснить связыванием с белком за счет водородных связей с GLN76(В), ассоциированной с белком молекулой воды и гидрофобной связи c LYS81(В),гидрофобной связи сахарофосфатного остова с Ser70(B) и LEU71(B).Аналогично, деформация Т7 происходит за счет водородной связи с ассоциированной с белком молекулой воды, водородной связи сахарофосфатного остова с ассоциированной молекулой воды и гидрофобной связи с SER37(B).В целом,связывание ДНК с белком может приводить к сильной деформации ДНК для приобретения наиболее энергетически выгодной структуры. - Исследование природы ДНК-белковых контактов .
Будем считать полярными атомами- кислород и азот, а неполярными - атомы углерода, фосфора и серы. Полярный контакт- ситуация, в которой расстояние между полярным атомом белка и полярным атомом ДНК меньше 3.5A. Аналогично, неполярным контактом будем считать пару неполярных атомов на расстоянии меньше 4.5A.
Исходный скрипт(для нового формата pdb) для выполнения упражнения ( взаимодействие со всем белком):my.def.Таблица. Контакты разного типа в комплексе 1MHD.pdb со всем белком
Контакты атомов белка с Полярные Неполярные Всего остатками 2'-дезоксирибозы 1 9 10 остатками фосфорной кислоты 9 2 11 остатками азотистых оснований со стороны большой бороздки 8 6 14 остатками азотистых оснований со стороны малой бороздки 0 0 0
Больше всего контактов возникает между атомами белка и остатками фосфорной кислоты,а также остатками азотистых оснований со стороны большой бороздки. Не возникает контактов между атомами малой бороздки ДНК и белком.Это объясняется тем,что связывание с атомами малой бороздки ДНК стерически менее выгодно, так как фактически малая бороздка менее доступна для связывания . - Получение популярной схемы ДНК-белковых контактов с помощью nucplot
Команда для получения схемы: nucplot 1MHD_old.pdb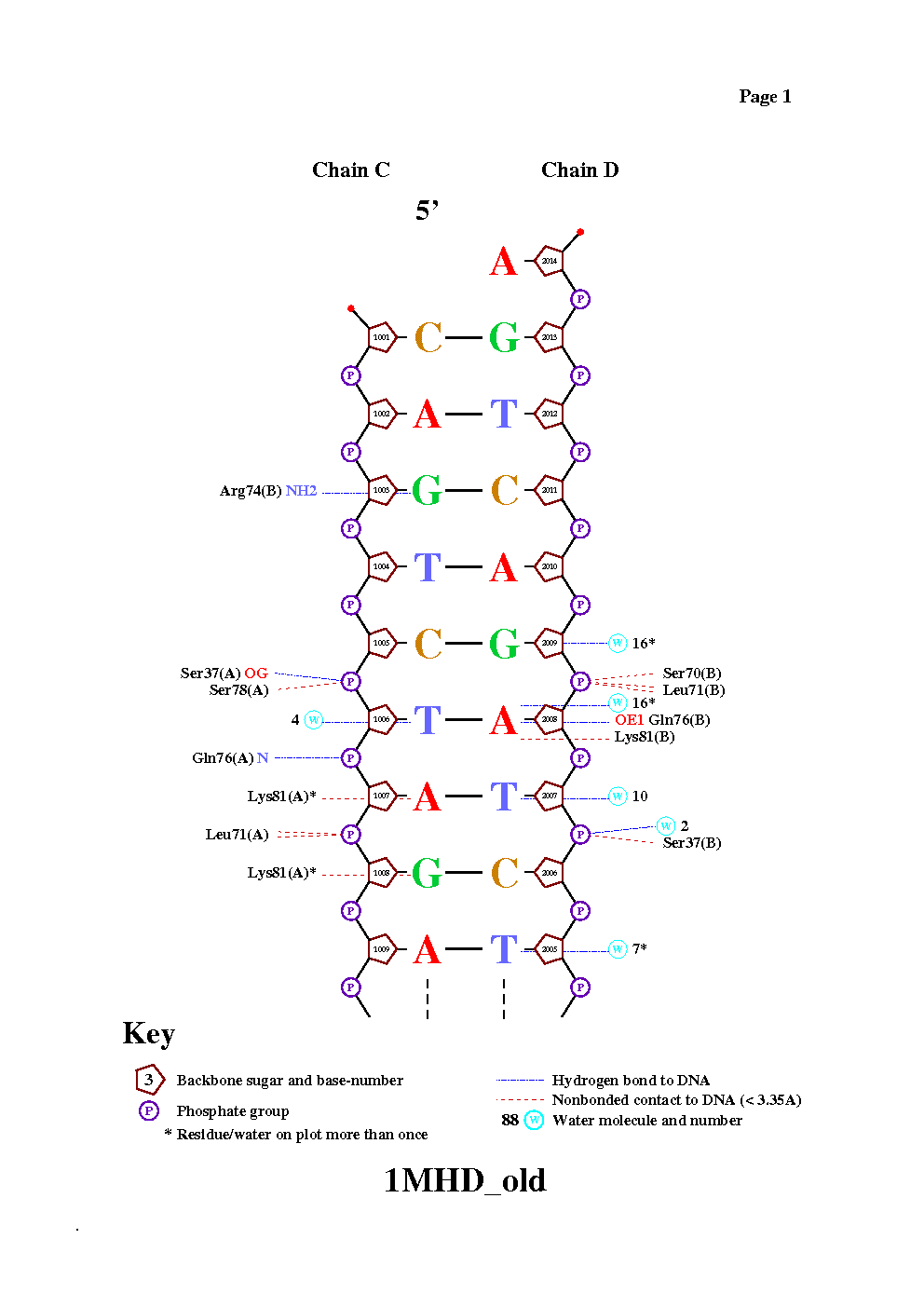
- Возможные распознающие контакты.
По моему мнению, наиболее важный аминокислотный остаток для распознавания последовательности ДНК -Arg74(B), так как он является распоспознающим нуклеотидом для гуанина и образует две водородные связи с ним, или GLN76(B),образующий водородную связь с А2008,являющийся предположительно распознающей аминоксилотой для аденина.
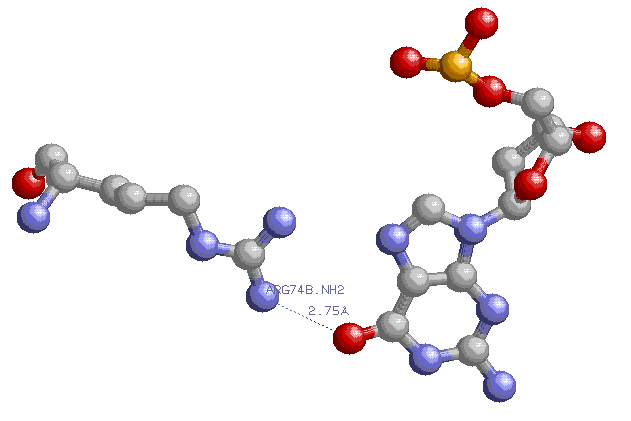
Контакт Arg74(B) и G1003.Водородная связь предположительно образована атомами NH2 аминокислоты и О6 гуанина.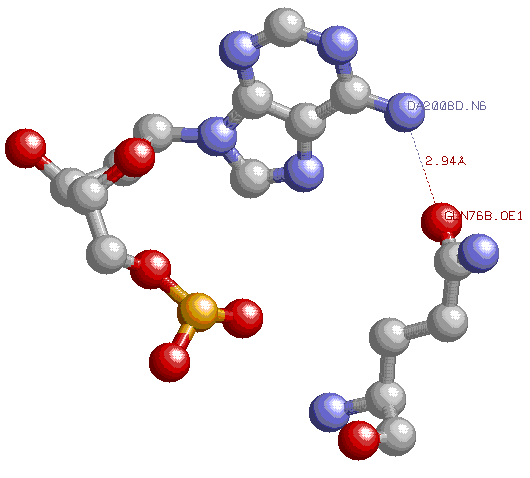
Контакт Gln76(B) и A2008.Водородная связь предположительно образована атомами OE1 аминокислоты и N6 аденина.
- Характеристика ДНК-связывающего домена MH1 (PF03165) при помощи базы данных Pfam.
Домен MH1( MAD-гомолог 1) расположен на N-конце родственных MAD белков,таких как белки из семейства SMAD. Этот домен отделен от домена MH2 неконсервативным связывающим регионом.Как показывает кристаллическая структура белка MH1, 11 высококонсервативных остатка бета-шпильки преданазначены для связывания с типичной GNCN последовательностью большой бороздки ДНК, необходимой для активации транскрипции нужных генов.Однако не все белки,содержащие этот домен,могут связываться с ДНК. Например,белок Smad2 не может связываться с ДНК и имеет длинную вставку между двумя последовательностями шпильки,что,вероятно,нарушает связывание с ДНК.Основная спираль (Н2) домена MH1 вместе сигналом ядерной локализации KKLKK являются основными факторами ядерного импорта белка SMAD3. Так же белки семества Smad используют домен MH1 для взаимодействия с факторами транскрипции, такими как Jun, TFE3, Sp1, и Runx [1,3].
В белках семества SMAD3 домен MH1 находится рядом с доменом MH2.
Cхематичное изображение расположения доменов.
Изображения доменов
На каждой картинке изображены белки,состоящие из двух одинаковых цепей, домены обозначены зеленым цветом .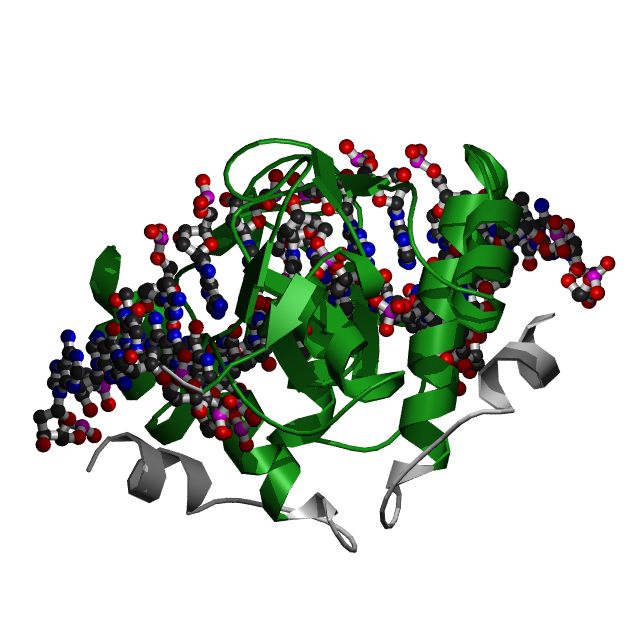
CRYSTAL STRUCTURE OF A SMAD MH1 DOMAIN BOUND TO DNA .(1mhd) CRYSTAL STRUCTURE OF SMAD3-MH1 BOUND TO DNA AT 2.4 A RESOLUTION .(1ozj) 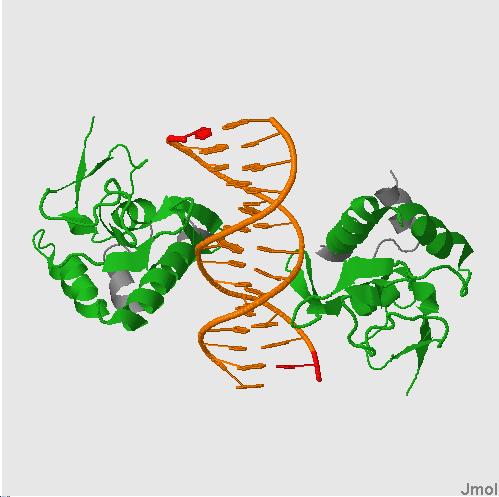
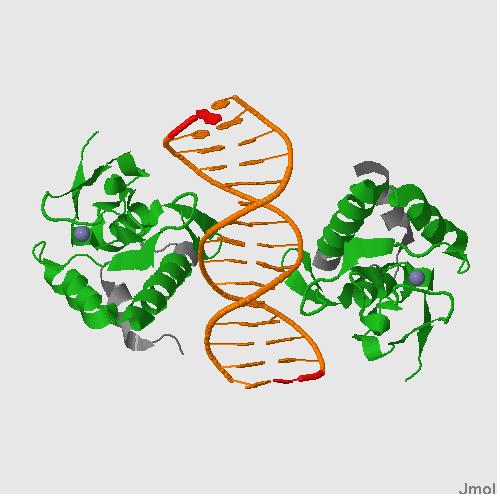
- Характеристика ДНК-связывающего домена MH1 (IPR013019 MAD homology, MH1) при помощи базы данных InterPro.
Домен MH1 имеет компактную глобулярную форму,состоящую из 4-х альфа-спиралей, 6 коротких бета-листов и 2 петель. N-концевая часть состоит из 3х альфа-спиралей, тогда как С-концевая часть содержит в себе 6 бета-тяжей,которые образуют две небольшие бета-пластины и одну бета-шпильку. Четвертая альфа-спираль находится в центральной части молекулы и окружена с одной стороны тремя альфа-спиралями N-концевой части,а с другой стороны двумя бета-пластинками и одной бета-шпилькой С-концевого участка. Эти дополнительные структурные элементы связаны с входящими в эту область петлями.
В домене MH1 находится ДНК-связывающий мотив из 11 аминокислотных остатков,расположенных на В2 и В3 бета-тяжах, для взаимодействия с большой бороздкой ДНК. Два остатка L3-петли и предшествующий им В2 бета-тяж также способствуют распознаванию последовательности ДНК.Бета-шпилька выступает наружу из центральной части домена.
Картинка со схематичным изображением доменов: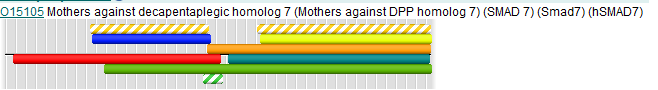
Количество различных контактов ДНК с молекулой белка, полученное в предыдущих упражнениях,не совпадает с таковыми,полученными при помощи программы nucplot. Это объясняется тем,что программа nucplot определяет взаимодействия между атомами и группами атомов более сложным способом,чем простой подсчет расстояния. В третьем упражнении при помощи подсчета торсионных углов в молекуле ДНК и при помощи визуализации наших результатов в программе Rasmol мы получили самые "деформированные" нуклеотиды двух цепей:10С и 12Т цепи С и 6А и 7Т цепи D. С помощью программы nucplot можно убедиться,что деформирование нуклеотидов 6А и 7Т цепи D происходит из-за связывания с ДНК, тогда как деформирование нуклеотидов 12Т и 10С цепи С не связано со связыванием с белком (на картинке они вообще не показаны).
Aминокислотный остаток с наибольшим числом указанных на схеме контактов с ДНК: LEU71 (Цепи А или В,так как они идентичны) образует две связи с атомом фосфора длиной меньше 3.35 ангстрем.
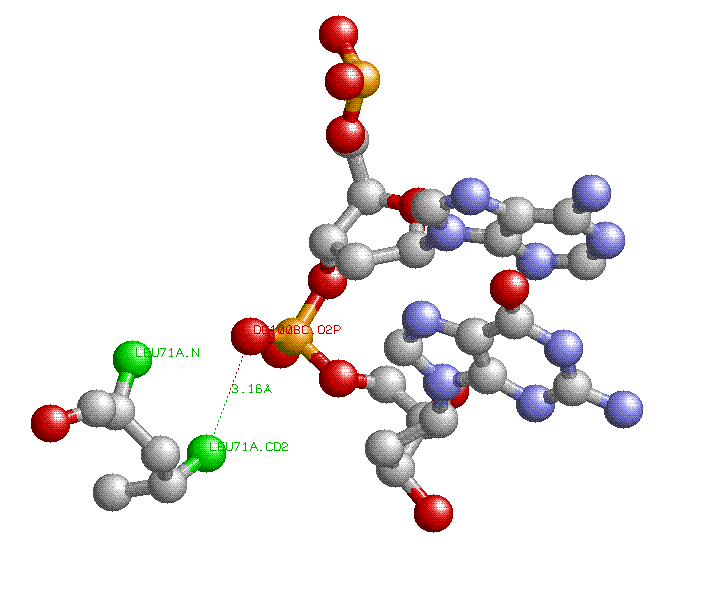
Примечание: на картинке зеленым показаны атомы Leu71:A, образующие связи с атомом кислорода DG1008C.O2P, длины этих связей 3.16 и 3.17 ангстрем(на картинке показана одна связь).