Исследование ДНК-белковых взаимодействий в структуре комплекса GCM-домена с ДНК
- Краткое описание структуры в файле 1ODH.pdb
- Функции белка, структура которого представлена в файле 1ODH.pdb
- Исследование структуры ДНК
- Исследование природы ДНК-белковых контактов
- Получение популярной схемы ДНК-белковых контактов с помощью nucplot
- Возможный(е) распознающий(е) контакт(ы)
- Характеристика ДНК-связывающего домена белка GCM1_MOUSE
В файле приведены координаты атомов следующих молекул: GCM-домен (из Mus musculus; 1 молекула), двухцепочечная ДНК (синтетическая; 2 олигонуклеотида, 2 молекулы). Для исследования были выбраны цепь A белка и цепи C и D, представляющие ДНК со следующей последовательностью:
цепь C [1001] 5' - cgatgcgggtgca - 3' [1013]
|||||||||||||
цепь D [1026] 3' - gctacgcccacgt - 5' [1014]
Функция белка GCM1_MOUSE: транскрипционный фактор, необходимый для развития плаценты (на основании схожести последовательности). Располагается в ядре клетки и является плацентоспецифичным. Содержит один GCM ДНК-связывающий домен.
ДНК представлена правозакрученной спиралью. Форма ДНК ближе всего к B-форме. Определение средних значений торсионных углов для внутренних нуклеотидов: ссылка на файл Excel. Самый "кривой" нуклеотид — 7-й гуанин цепи I (DG1007 цепи C).
Как видно, значения торсионных углов в данной связанной с белком ДНК значительно отклоняются как от их среднего значения, так и от значений, характерных для "типичной" B-ДНК (настолько, что для некоторых пар нуклеотидов (5-я - 10-я) analyze не указывает наиболее близкий тип формы ДНК). Отсюда понятно, что связывание с белком приводит к достаточно заметной деформации.
| Контакты атомов белка с | Полярные | Неполярные | Всего |
| остатками 2'-дезоксирибозы | 2 | 9 | 11 |
| остатками фосфорной кислоты | 12 | 8 | 20 |
| остатками азотистых оснований со стороны большой бороздки | 4 | 9 | 13 |
| остатками азотистых оснований со стороны малой бороздки | 0 | 0 | 0 |
Скрипт, определяющий множества атомов:
define set1 (dna and *.O?*)
define set2 (dna and *.O?P)
define set3 (dna and *.N?)
Остатки фосфорной кислоты содержат относительно много полярных атомов (кислорода), обращенных наружу, поэтому образуют много полярных контактов. Полярные атомы остатков 2'-дезоксирибозы и азотистых оснований уже расположены "глубже", поэтому образуют меньше полярных контактов. Остатки азотистых оснований, как видно, образуют контакты только со стороны большой бороздки. Это показывает, что специфичное связывание белка с какой-либо определенной последовательностью ДНК обусловлено образованием контактов между азотистыми основаниями и аминокислотными остатками именно в большой бороздке.
Использованная команда: nucplot 1ODH_old.pdb
Полученная картинка:
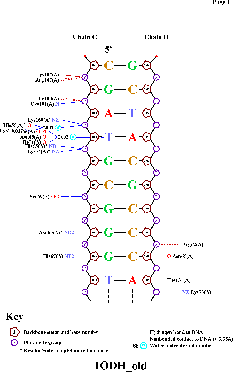
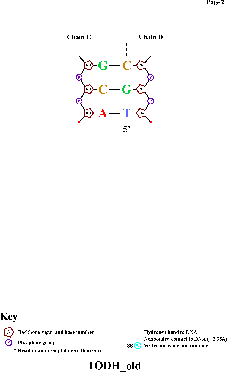
Полученная картинка показывает, что аминокислотные остатки образуют контакты именно с теми нуклеотидами, у которых (а также у образующих с ними комплементарные пары нуклеотидов) торсионные углы наиболее значительно отличаются от средних (пример — 4-я пара нуклеотидов).
Наибольшее число контактов между аминокислотным остатком и ДНК, согласно схеме, — 2. Это остатки His55, Asn65 и Lys74, однако один из контактов His55 и Lys74 образуют через молекулу воды, тогда как Asn65 образует оба контакта непосредственно с ДНК. Поэтому аминокислотный остаток с наибольшим числом указанных на схеме контактов с ДНК — Asn65.
По моему мнению, контакты остатка Asn65 являются наиболее важными для распознавания последовательности ДНК:
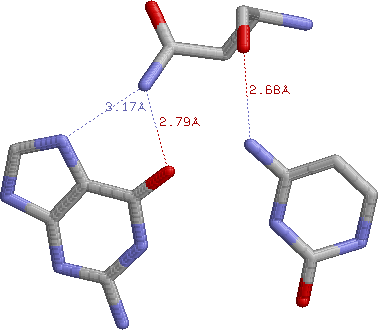
Этот остаток образует 3 явные водородные связи с двумя азотистыми основаниями (причем они принадлежат двум нуклеотидам разных цепей, не являющимся комплементарной парой, т.е. Asn65 распознает сразу две соседние пары оснований):
Pfam показывает присутствие в белке двух доменов: GCM-домена (Glia cell missing; ДНК-связывающий домен) и домена, сгенерированного автоматическим выравниванием. Доменная структура:

GCM-домен (IPR003902 Tscrpt_reg_GCM_motif) обнаружен в белках, принадлежащих семейству регуляторов транскрипции и представляющих собой GCM-транскрипционные факторы Drosophila melanogaster и их гомологов у млекопитающих. Он составляет N-концевую часть GCM-транскрипционных факторов, тогда как C-концевая часть менее консервативна. Показано, что GCM-домен представляет собой ДНК-связывающий домен, распознающий преимущественно последовательность 5'-ATGCGGGT-3'. Он содержит много консервативных основных аминокислотных остатков, 7 остатков цистеина и 4 остатка гистидина. Цистеины участвуют в определении общей конформации домена, в процессе связывания ДНК и redox-регуляции этого связывания. GCM-домен представляет новый класс Zn-содержащих ДНК-связывающих доменов, непохожих на другие ДНК-связывающие домены.