Вторичная структура тРНК и ДНК-белковые контакты
Задание 1. Предсказание вторичной структуры заданной тРНК
Упражнение 1
С помощью программы einverted из пакета EMBOSS была предсказана вторичная структура глициновой тРНК. При всевозможных значениях параметров был предсказан только акцепторный стебель, причём в нём было предсказано на одну пару оснований больше, чем в реальности. Это объясняется тем, что в fasta-файле, подаваемом на вход einverted, присутствуют дополнительные 5 концевых нуклеотидов, которых в pdb-файле нет.
Упражнение 2
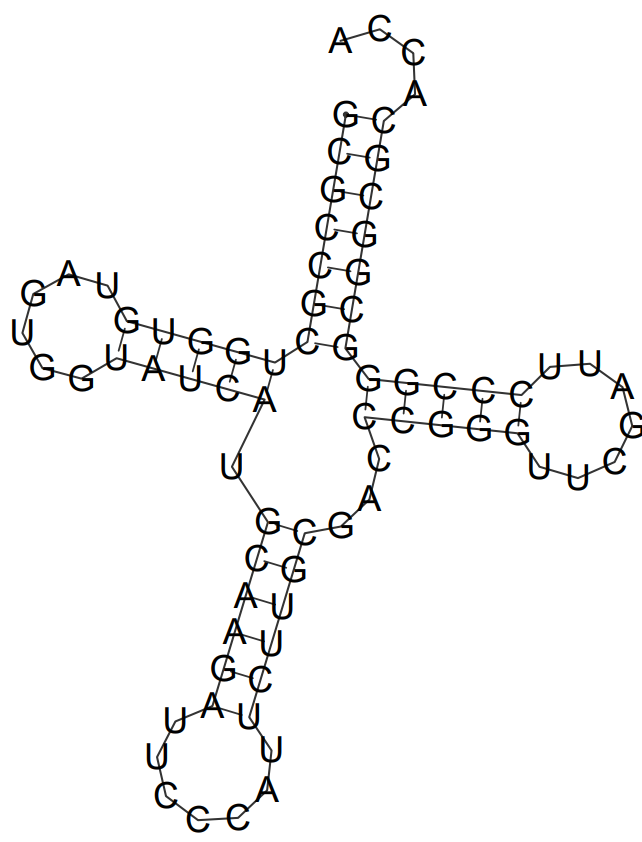
| Позиции в структуре (по результатам find_pair) | Результаты предсказания с помощью einverted | Результаты предсказания по алгоритму Зукера | |
|---|---|---|---|
| Акцепторный стебель | 5'-460-465-3'
5'-522-527-3' Всего 6 пар |
предсказано 6 пар из 6 реальных и 1 дополнительная | предсказано 6 пар из 6 реальных и 1 дополнительная |
| D-стебель | 5'-468-471-3'
5'-479-482-3' Всего 4 пары |
- | предсказано 0 пары из 4 реальных и 5 дополнительных (цепи сдвинуты друг относительно друга на 3 нуклеотида по сравнению с моделью find_pair) |
| T-стебель | 5'-505-509-3'
5'-517-521-3' Всего 5 пар |
- | предсказано 6 пар из 7 реальных |
| Антикодоновый стебель | 5'-495-501-3'
5'-483-489-3' Всего 7 пар |
- | предсказано 5 пар из 5 реальных |
| Общее число канонических пар нуклеотидов | 18 | 7 | 21 |
Задание 2. Поиск ДНК-белковых контактов в заданной структуре
В качестве ДНК, связанной с белком, была выбрана ДНК, связанная с транскрипционным фактором PAP1 (PDB ID: 1GD2).
Упражнение 1
Для запуска в Jmol был создан скрипт, задающий множества атомов, и скрипт, показывающий описанные в задании структуры.
Упражнение 2
Количество контактов каждого типа (Табл. 2) было получено с помощью скрипта contacts.spt - при запуске в консоли команда select выдаёт количества контактирующих с ДНК атомов белка. Оказалось, что ДНК образует с лейциновой молнией белка PAP1 в 2 раза больше неполярных контактов, чем полярных. Стоит отметить, что остатки азотистых оснований контактируют с белком почти исключительно через большую бороздку, а атомы кислорода остатков 2'-дезоксирибозы практически не образуют контактов.
| Полярные | Неполярные | Всего | |||
|---|---|---|---|---|---|
| Остатками 2'-дезоксирибозы | 4 | 46 | 50 | ||
| Остатками фосфорной кислоты | 22 | 15 | 37 | ||
| Остатками азотистых оснований со стороны большой бороздки | 29 | 44 | 73 | ||
| Остатками азотистых оснований со стороны малой бороздки | 2 | 6 | 8 | ||
Упражнение 3
С помощью программы nucplot была получена схема ДНК-белковых контактов. Она показывает контакты с четырьмя, а не двумя цепями, из-за того, что PAP1 взаимодействует с двумя изолированными участками ДНК. Так как взаимодействия с обоими участками симметричны, в следующем упражнении рассматривается только одно из них (с цепями A и B).
Упражнение 4
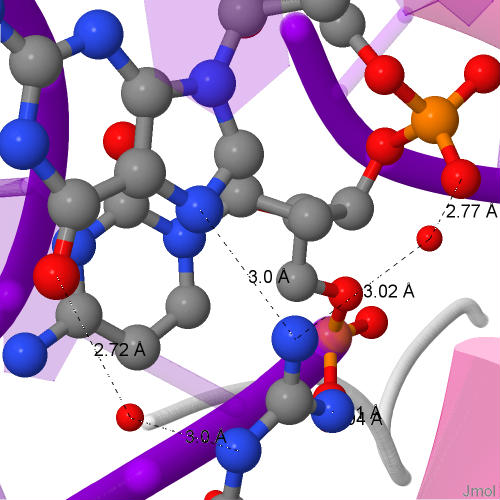
Все взаимодействующие с ДНК остатки непосредственно образуют с ней только по 1 контакту. Но если учесть контакты через молекулы воды, то больше всего контактов образуют остатки Arg94 на E и F цепях PAP1 (показано для цепи только F). Они же являются самыми важными для распознавания последовательности ДНК, так как распознают гуанин, контактируя через молекулу воды с его атомом O6 и непосредственно атомом N7 [1].
Список литературы
- S.S. Chavali, C.E. Cavender, D.H. Mathews, and J.E. Wedekind. Arginine Forks Are a Widespread Motif to Recognize Phosphate Backbones and Guanine Nucleobases in the RNA Major Groove. J. Am. Chem. Soc. 2020, 142, 47, 19835–19839. DOI: 10.1021/jacs.0c09689