Данные по геному бактерии Deinococus radiodurans R1
Файлы можно скачать по ссылке ниже:
- Excel-файл с информацией о геноме в целом.
- Excel-файл с информацией о квазиоперонах и пересечениях генов.
Краткие обозначения, используемые в файле и ниже:
- Сhr1 - первая хромосома.
- Chr2 - вторая хромосома.
- MP1 - плазмида MP1.
- CP1 - плазмида CP1.
- CDS - совокупность белковых генов участка ДНК.
- RNA - совокупность РНКовых генов участка ДНК.
Таблица 1
Количество квазиоперонов в хромосомах и плазмидах (пороговое расстояние 100, по стандарту). Я считал гены пересекающимися, если координаты конца предыдущего гены совпадали или были больше, чем координаты начала следующего. Рассматривались только белковые гены. | ||||||||||||||||||||||||||||||||||||
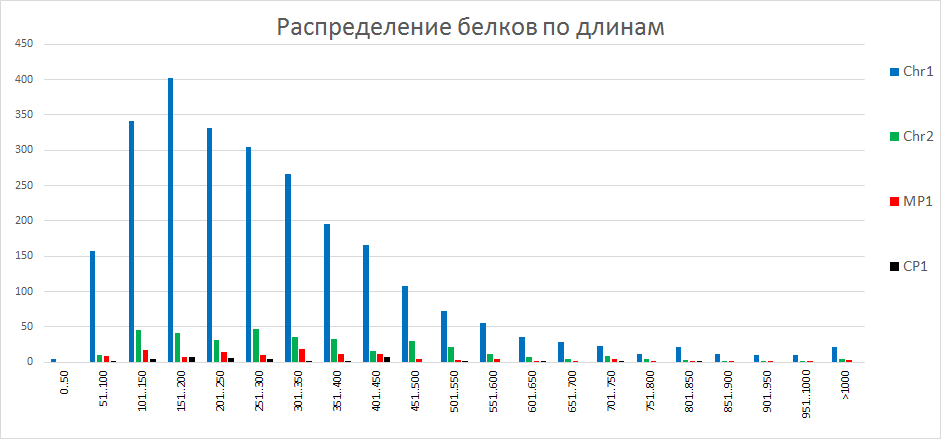
| Рисунок 1
На гистограмме представлено распределение белков различных участков ДНК по длинам. Из этого графика видно, что наибольшее количество белков сосредоточено в хромосоме 1. Наибольшее количество белков сосредоточено в диапазоне длины от 101 до 350 аминокислот. | ||||||||||||||||||||||||||||||||||||
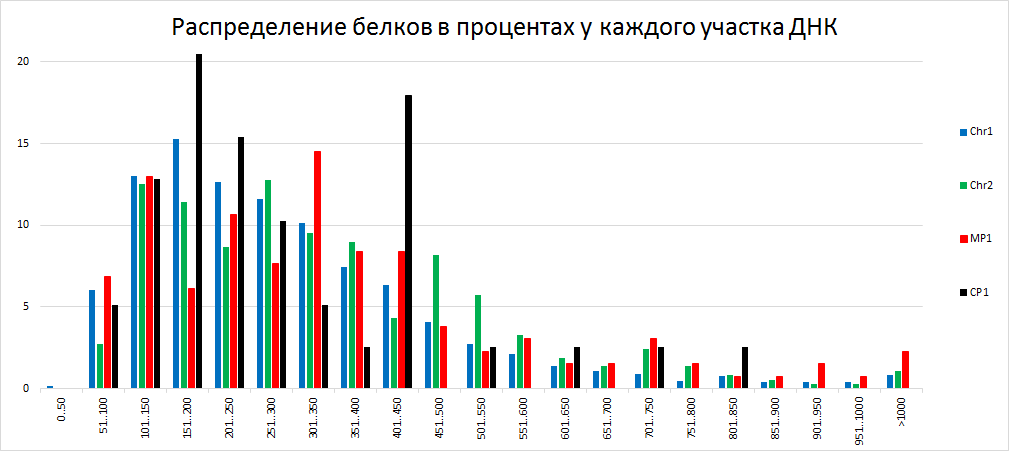
| Рисунок 2
Гистограмма процентного распределения белков по длинам для каждого участка ДНК позволяет понять, белки какой длины преобладают в той или иной хромосоме или плазмиде. Так например, плазмида MP1 имеет большое количество белков высокой длины (>901). А у плазмиды CP1 есть 5 диапазонов длин, содержащих наибольшее количество белков (>10), при этом по одному из них процентное содержание больше 20 процентов. | ||||||||||||||||||||||||||||||||||||
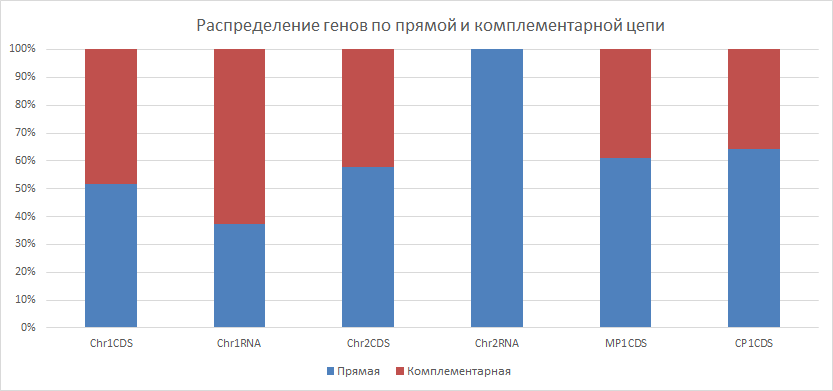
| Рисунок 3
Из гистограммы видно, что гены преимущественно располагаются на прямой цепи, и только для РНК генов первой хромосомы это не выполнятеся. Стопроцентный показатель у РНК генов второй хромосомы объясняется невысоким общим количеством генов (всего 1 ген). | ||||||||||||||||||||||||||||||||||||
| Рисунок 4
Из гистограммы видно, что наибольшее количество генов имеет длину кратную 3 (>95%). Однако, если мы посмотрим отдельно на белковые и РНКовые гены, то увидим огромную разницу: для белковых генов характерна делимость на 3, а для РНКовых нет (<20% для РНК генов хромосомы 1). О возможных причинах этого поговорим позже. | ||||||||||||||||||||||||||||||||||||
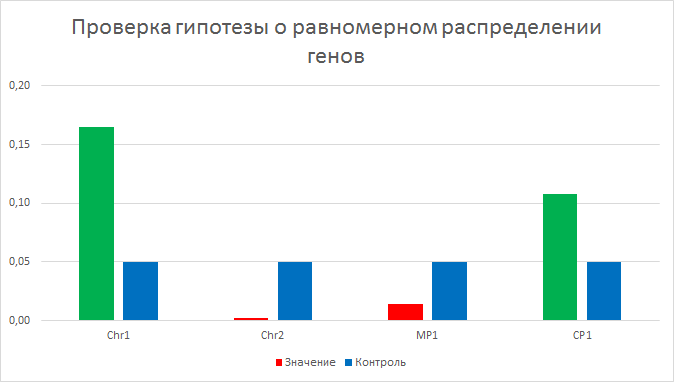
| Рисунок 5
Гипотезу о том, что гены распределены случайно с вероятностью 0.5 проверили с помощтю биномиального распределения (все формулы в файле). Рассматривались только белковые гены. Реузльтаты видны в гистограмме, где синим цветом обозначены контрольные значения в 0.05. Все значения выше контрольного считаются подтверждающими гипотезу и обозначены зеленым, все значения ниже - красным. Гипотеза подтвердилась для первой хромосомы и плазмиды CP1. | ||||||||||||||||||||||||||||||||||||
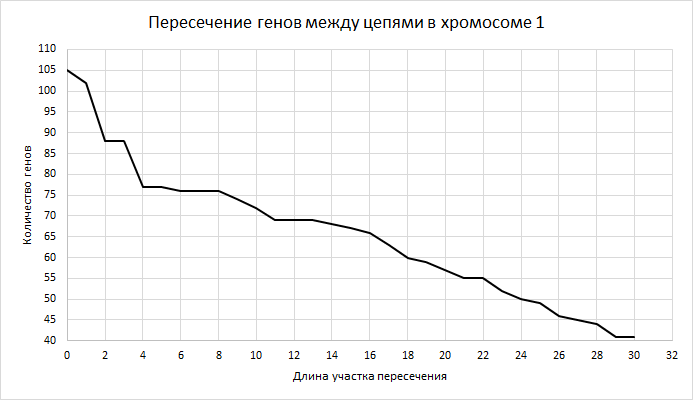
| Рисунок 6
На графике показано пересечение генов между комплементарными цепями в хромосоме 1. Здесь и далее по оси Х отложена длина участка пересечения, причем нулю соответствует пересечение на 1 нуклеотид, так как расчет проводился по формуле координаты конца минус координаты начала. По оси У - количество генов с таким пересечением. Значение быстро падает от длины пересечения в 3 (4 нуклеотида) к 4, а потом плавно снижается. Далее 30 не рассматривалось. | ||||||||||||||||||||||||||||||||||||
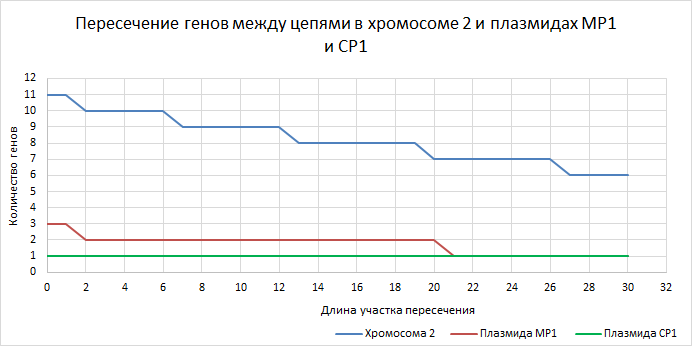
| Рисунок 7
Для хромосомы 2 и плазмид количество пересечений невелико. Их количество плавно снижается. | ||||||||||||||||||||||||||||||||||||
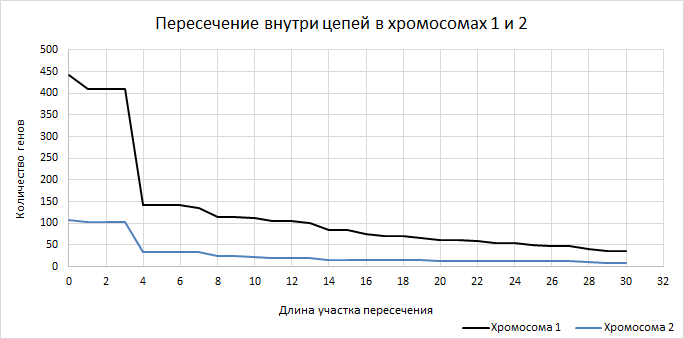
| Рисунок 8
Пересечение генов внутри цепей у хромосом показывает очень интересную закономерность: после длины пересечения в 3 количество пересекающихся генов резко падает, а потом изменяется незначительно. | ||||||||||||||||||||||||||||||||||||
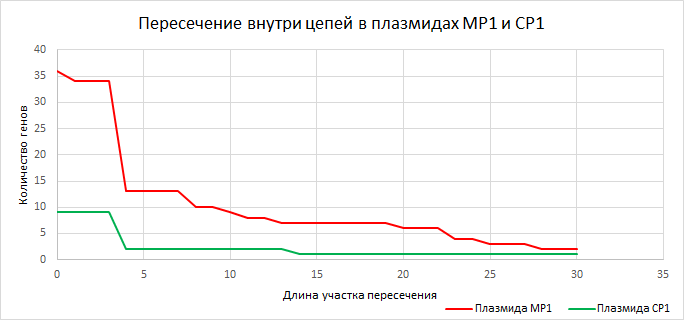
| Рисунок 9
Закономерность, выявленная для хромосом подтверждается и для плазмид. | ||||||||||||||||||||||||||||||||||||
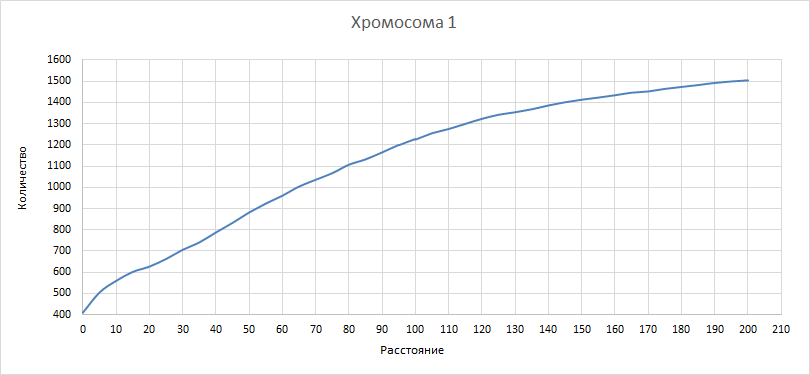
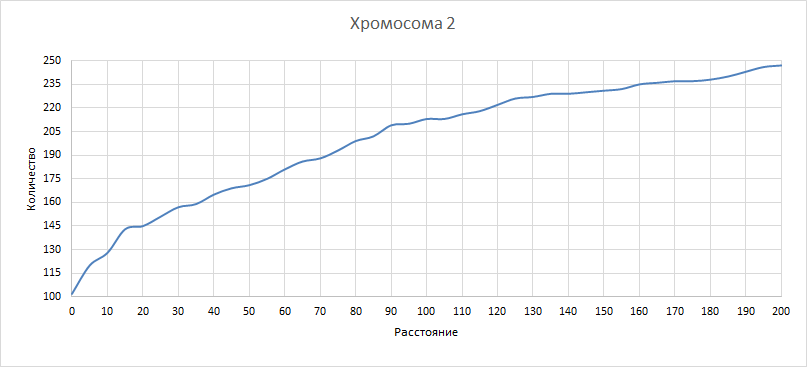
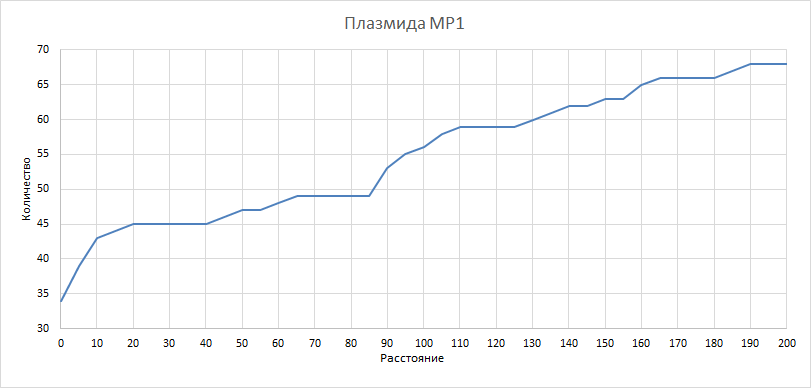
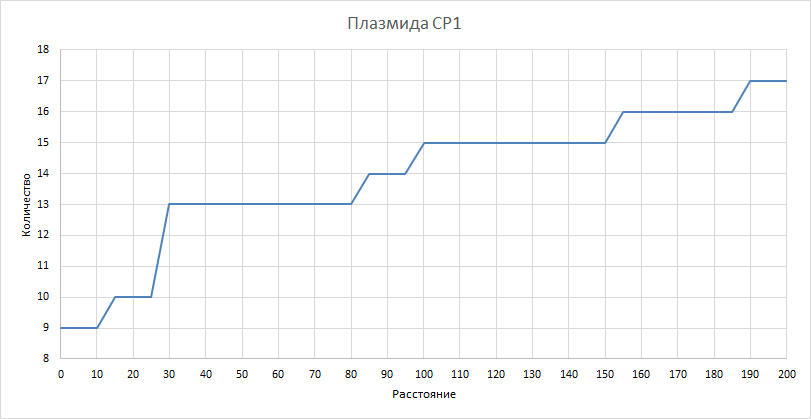
| Рисунок 10
Здесь и в последующих трех графиках рассматривали, как изменилось бы количество квазиоперонов при изменении расстояния порога. Рассматривали значения порога от 0 до 200 с шагом 5. Как и ожидалось, количество квазиоперонов падает при уменьшении порога и увеличивается при его увеличении. | ||||||||||||||||||||||||||||||||||||
| Рисунок 12
Наблюдаем все сказанное выше для хромосомы 1. | ||||||||||||||||||||||||||||||||||||
| Рисунок 13
Наблюдаем все сказанное выше для хромосомы 1. | ||||||||||||||||||||||||||||||||||||
| Рисунок 14
Наблюдаем все сказанное выше для хромосомы 1. |
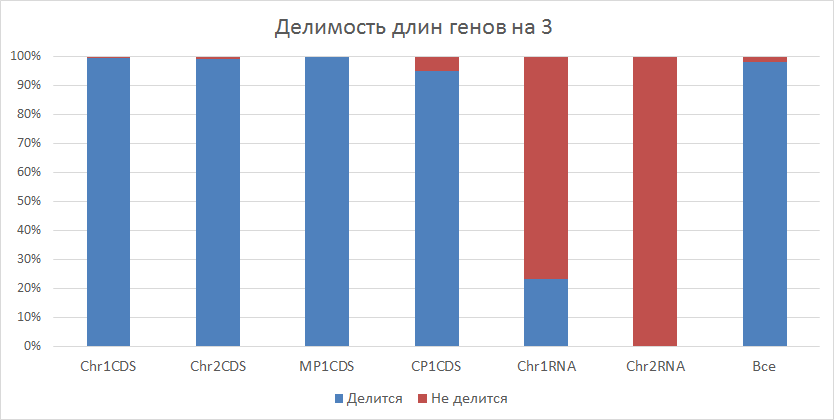
Из рисунка 4 видно, что практически все гены, кодирующие белки, имеют длину кратную 3, и большинство
генов, кодирующих РНК, не имеют данного свойства. Я считаю, что основная причина этого - интроны.
Интрон - любая последовательность нуклеотидов в гене, которая удаляется в процессе слайсинга
и не содержится в финальной цепи РНК. Интроны найдены в генах большинства организмов и многих вирусов.
Интроны встречаются в генах, кодирующих рРНК, тРНК и белки. Длина интронов может не делиться на 3.
Хотя интроны и характерны в основном для эукариот, но прокариоты тоже имеют интроны. Интроны прокариот
вырезаются в процессе автослайсинга. Исходя из рисунка 4 можно судить, что интроны практически не
характеры для белков, однако часто встречаются у РНКовых генов.