Предсказание вторичной структуры тРНК. Поиск ДНК-белковых контактов.
Задание 1. Предсказание вторичной структуры тРНК разыми алгоритмами.
По данным работы функции einverted и алгоритма
Зукера получил 2 предсказания вторичной структуры. Данные занесены в
таблицу 1. Обсуждение работы алгоритмов ниже таблицы.
| Таблица 1. Реальная и предсказанная вторичная структура тРНК из файла 1o0b.pdb | |||
|---|---|---|---|
| Участок структуры | Позиции в структуре | Результаты предсказания с помощью einverted | Результаты предсказания по алгоритму Зукера |
| Акцепторный | 5'-902-907-3' 5'-966-971-3' Всего 6 пар | 5'-901-907-3' 5'-965-971-3' Предсказано 6 пар из 6 + 1 лишняя | 5'-901-907-3' 5'-964-970-3' Предсказано 6 пар + лишняя, но произошло смещение по второй цепи |
| D-стебель | 5'-910-912-3' 5'-923-925-3' Всего 3 пар | - | 5'-910-912-3' 5'-922-924-3' Предсказано 3 пары, произошло смещение на 1 по второй цепи |
| T-стебель | 5'-949-953-3' 5'-961-965-3' Всего 5 пар | - | 5'-947-951-3' 5'-959-963-3' Предсказано 5 пар, смещение по всем осям на 2 |
| Антикодоновый стебель | 5'-926-933-3' 5'-937-944-3' Всего 8 пар | - | 5'-926-930-3' 5'-938-941-3' Предсказано 5 пар из 8 реальных |
| Общее число канонических пар | 28 пар | 7 пар | 20 пар |
Единственная осмысленная выдача программы einverted имеет счет 21, при уменьшении минимального счета до очень низких значения программа не дает осмысленных результатов. Однако, программа, работающая по алгоритму Зукера, показала очень хорошие результаты. С высокой точностью была предсказана вторичная структура тРНК. Кроме того, программа обладает граической выдачей (рисунок ниже).
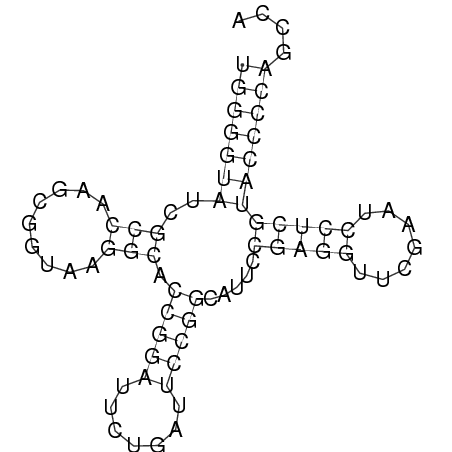
Задание 2. ДНК-белковые контакты.
В ходе выполнения упражнений этого задания написал
скрипты, все ссылки представлены далее. Для определения множества
атомов написал definition.spt , для
последовательного вывода с паузами написал
imagines.spt , для определения числа
контактов - contacts.spt . Так как для
задания мне необходима только A цепь белка, создал с помощью JMol
обрезанную версию файла 1p47.pdb -
1p47m.pdb, которая содержит только ДНК и
А цепь белка. Все скрипты писались для этого файла. Данные по контактам
представлены в таблице 2.
| Таблица 2. Контакты разного типа в комплексе 1p47m.pdb (только А цепь белка) | |||
|---|---|---|---|
| Контакты атомов белка с | Полярные | Неполярные | Всего |
| остатками 2'-дезоксирибозы | 1 | 13 | 14 |
| остатками фосфорной кислоты | 11 | 10 | 21 |
| остатками азотистых оснований со стороны большой бороздки | 13 | 25 | 38 |
| остатками азотистых оснований со стороны малой бороздки | 0 | 4 | 4 |
Подсчет количества контактов проводился следующим образом: искалось количество атомов в белке, способных вступить в контакт с ДНК, и наоборот, из двух множеств выбиралось наибольшее. Количество членов этого множества считал количеством контактов, делал так, потому что один атом может образовать несколько контактов, но маловероятно, что атом из большего множества будет образовывать нескольно связей с атомами из меньшего множества.
Подведем некоторые итоги из данных таблицы:
1.Неполярные контакты преобладают над полярыми во всех группах, кроме фосфорной кислоты. Во-первых, критерий образования неполярной связи ниже, во-вторых, количество неполярных атомов во всех группах, кроме фосфорной кислоты, больше количества полярных;
2. Через фосфат образуется большее количество контактов, чем через сахар. Фосфат имеет более внешнее положение, нежели сахар, поэтому образовывать связи ему проще, несмотря не намного меньшее количество атомов.
3. Через большую бороздку образуется почти в 10 раз больше контактов, чем через малую. Во-первых, в В-форме в сторону большой бороздки смотрит большинство атомов оснований. Во-вторых в большой бороздке больше места для размещения множества аминокислотных остатков белка. В-третьих (предположение), большая бороздка играет большую роль в поддержании формы ДНК в ядре.
Получил схему ДНК-белковых контактов с помощью nucplot. Представлена только картинка, где расположены большинство контактов А цепи белка с ДНК.
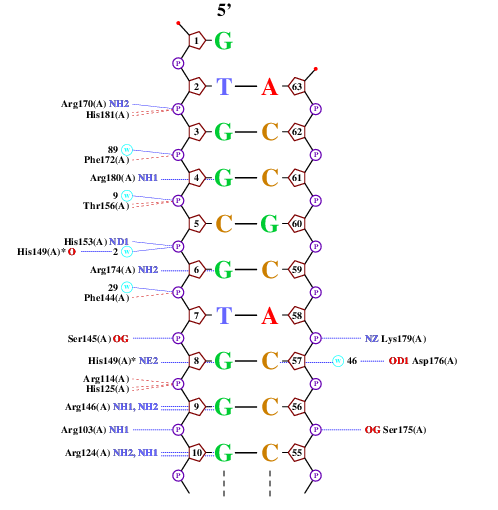
На данной схеме выбрал аминокислотных остаток, образующий наибольшее (в данном случае - 2) количество связей. Это Arg146(A). Аминокислотных остаток, наиболее важный для распознавания последовательности ДНК - тот, который связывается в азотистым основанием. Многие аргинины и гистидины на схеме связаны с одним видом азотистых оснований - гуанином. Возьмем, например, His149(A).
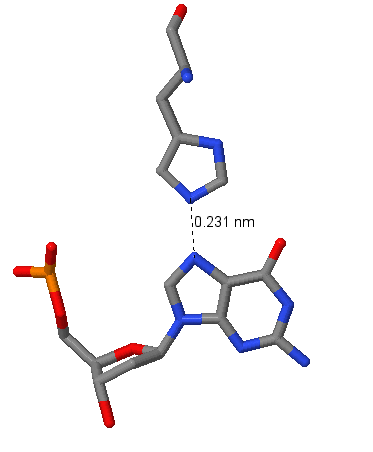
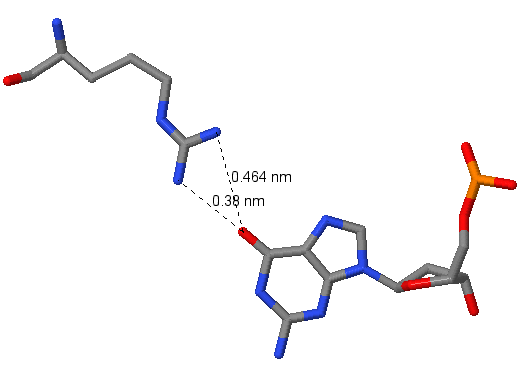 | 
|
| Рис.3. Аргинин в связи с гуанином | Рис.4. Гистидин в связи с гуанином |
На картинках видим контакты положительно заряженных аминокислот с азотистым основанием гуанином. Связь проходит по донорно-акцепторному механизму, т.е. положительно заряженный атом водорода (протон) с пустой s-орбиталью "берет" у атома-донора 2 электрона с р-орбитали. Это механизм образования водородной свзяи. Атомы водорода (мы их не видим), свзяанные с N в аргинине, берут электронную плотность у O, связанного с C двойной связью и имеющего 3 заполненных р-орбитали, который может образовать таким образом 3 водородные связи. В случае гистидина водородная связь образуется с протоном, присоединенным к имидазольному кольцу.
© Maximov Vladislav, 2019.