Восстановление кристалла из PDB файла
Для работы была выбрана ненасыщенная рамногалактуронил-гидролаза Bacillus subtilis (PDB 2D8L), используемая в предыдущих практикумах. Разрешение ее структуры составляет 1.7 А. Кристаллографические характеристики указаны в поле CRYST1 в файле PDB:
| Длины направляющих векторов | Углы между направляющими векторами | Кристаллографическая группа | Число молекул в ячейке | |
| CRYST1 | 91.831, 91.831, 178.865 | 90.00, 90.00, 120.00 | P 61 2 2 | 12 |
Элементарная ячейка 2D8L относится к кристаллографической группе P, то есть имеет примитивную центровку, и является гексагональной (α=β≠γ, α=β=90°, γ=120°).
Белок 2D8L является ассиметричным и состоит из одной цепи А (рис.1).
Рис 1. Структура 2D8L.
Для того, чтобы восстановить кристаллографические ячейки, использовалась следующая команда:
symexp sym, 2D8L, 2D8L, 3
Получилось, что на расстоянии менее чем 3Å от исходной структуры располагаются 8 ячеек (рис.2).
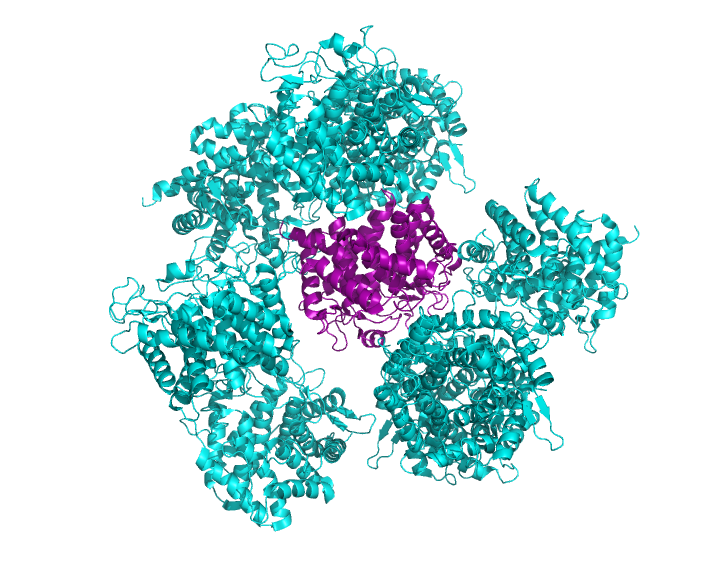
Рис 2. Структура 2D8L с восстановленными соседними ячейками, находящимися на расстоянии не более 3Å.
Для исследования контактов между молекулами в кристалле были отобраны исходная структура и один ее сосед, который впоследствии был переименован в cell. Для того, чтобы показать водородные связи между белками были созданы два множества остатков, находящихся на расстоянии 3,5Å от каждой структуры, но входящие в состав соседнего белка:
create ambience1, byres (2d8l within 3.5 of cell)
create ambience2, byres (cell within 3.5 of 2d8l)
distance hbonds, ambience1, ambience2, 3.5, 2
В последней команде distance был использован последний параметр со значением 2, который позволяет получить данные о полярных взаимодействиях. Таким образом, была получена картинка (рис.3) с большим количеством расстояний между аминокислотами, но при подробном изучении были отобраны наиболее подходящие для образования водородных связей (рис.4).
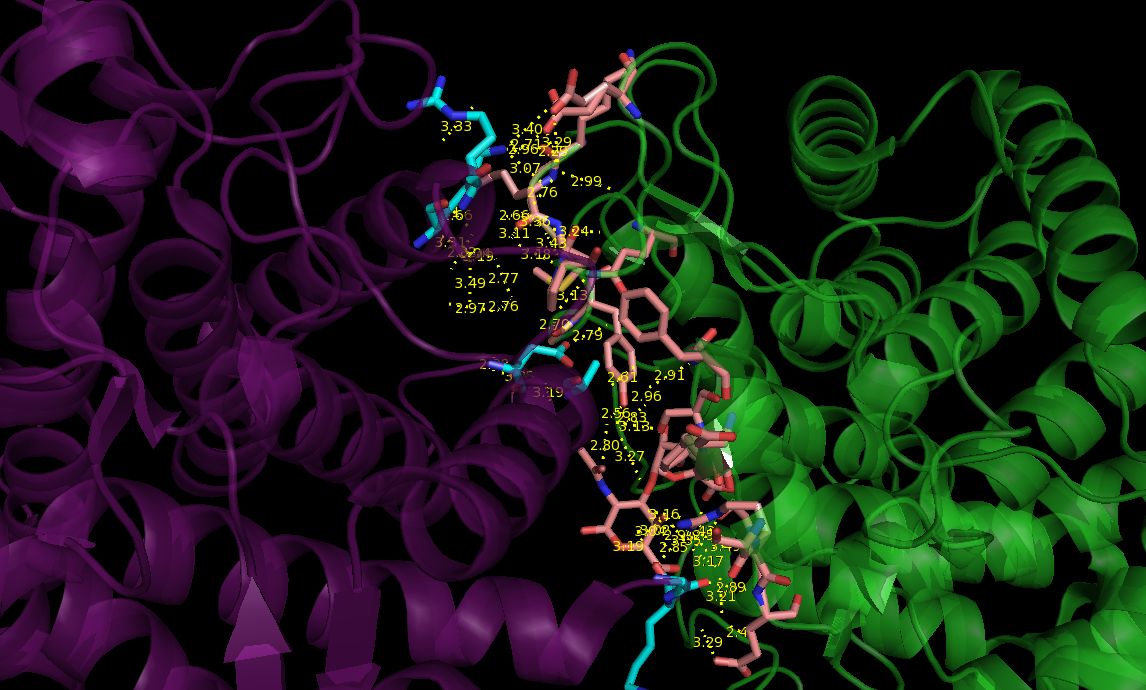
Рис 3. Контакты между 2D8L и соседней ячейкой, находящимися на расстоянии не более 3.5Å (фиолетовым обозначена структура 2D8L, зеленым - соседняя ячейка; остатки голубого цвета входят в состав 2D8L, а розового - в соседний белок).
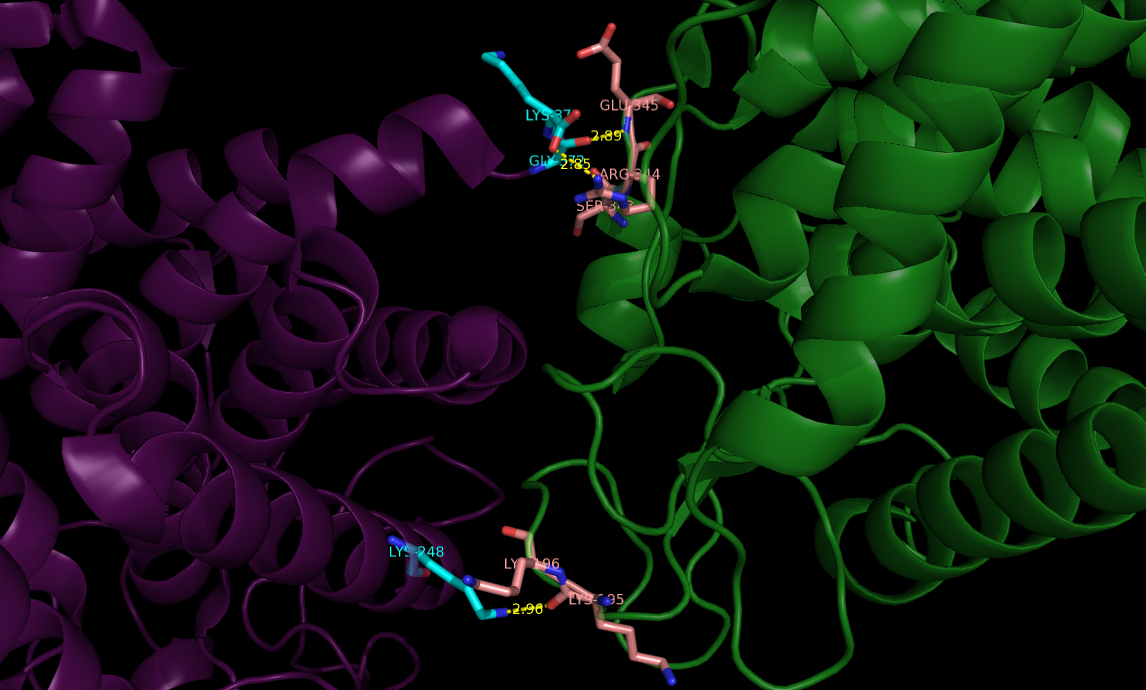
Рис 4. Водородные связи между остатками 2D8L и соседней ячейкой, находящимися на расстоянии не более 3.5Å (фиолетовым обозначена структура 2D8L, зеленым - соседняя ячейка; остатки голубого цвета входят в состав 2D8L, а розового - в соседний белок).
Далее были рассмотрены гидрофобные взаимодействия. С помощью следующей команды были выделены все неполярные аминокислоты (рис.5) в исходной структуре и соседней ячейке:
create hydph, resn ala+met+trp+phe+val+ile+leu
create hydph_2d8l, 2d8l and resn ala+met+trp+phe+val+ile+leu
create hydph_cell, cell and resn ala+met+trp+phe+val+ile+leu
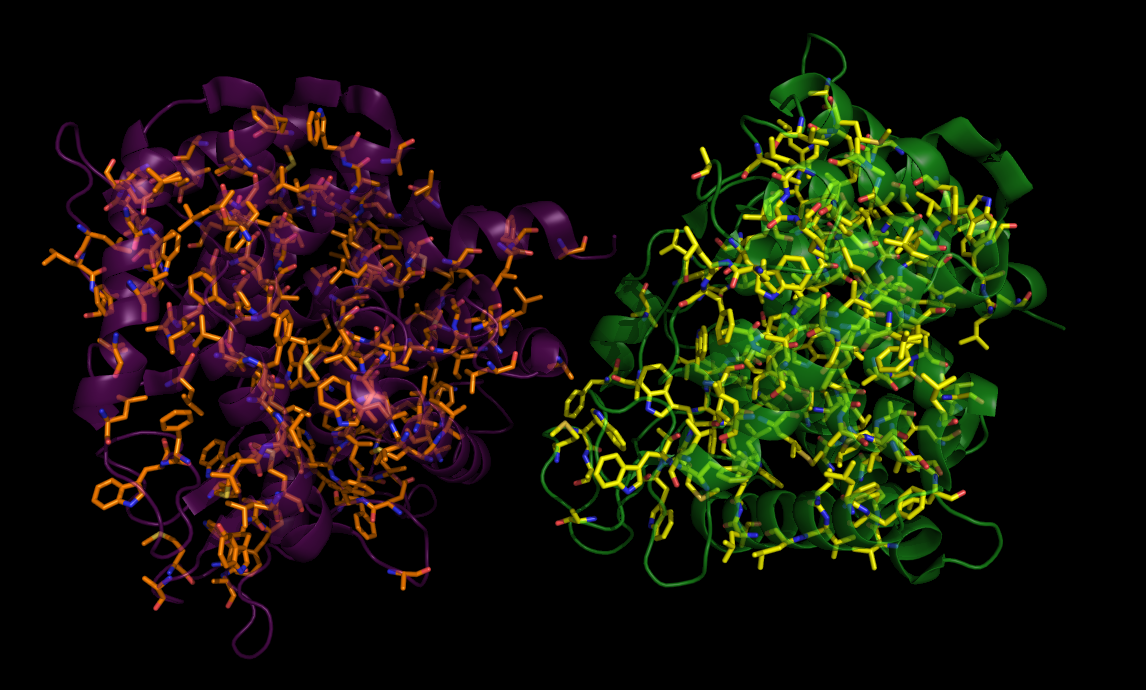
Рис 5. Гидрофобные остатки (обозначены оранжевым и желтым цветом) в 2D8L и соседней ячейке (фиолетовым обозначена структура 2D8L, зеленым - соседняя ячейка).
Далее были выделены неполярные остатки обеих структур, находящихся на расстоянии не более 4-х ангстрем (рис.6). Таковых оказалось три, предполагается, что между ними возникают гидрофобные взаимодействия.
create ambience3, byres (hydph_2d8l within 4 of hydph_cell)
create ambience4, byres (hydph_cell within 4 of hydph_2d8l)

Рис 6. Остатки (обозначены оранжевым и желтым цветом) в 2D8L и соседней ячейке, между которыми возникают гидрофобные взаимодействия (фиолетовым обозначена структура 2D8L, зеленым - соседняя ячейка).
Таким образом,по всей видимости, водородные связи играют большую роль в кристаллизации белка.
Ссылка на проект в PyMol: pr2.pse
Для дальнейшей работы была выбрана структура 3HDD - комплекс гомеодоменового белка с ДНК Drosophila melanogaster, имеющая разрешение 2.2 ангстрем.
Для построения соседних молекул ДНК, которая контактирует с цепью B белка (не дальше, чем на расстоянии 4-х ангстрем), расположенной "на краю" ДНК, была использована следующа команда:
symexp sym, 3hdd, chain B, 4
На рис.7 и рис.8 видно, что цепь В контактирует с молекул ДНК из соседней ячейки.
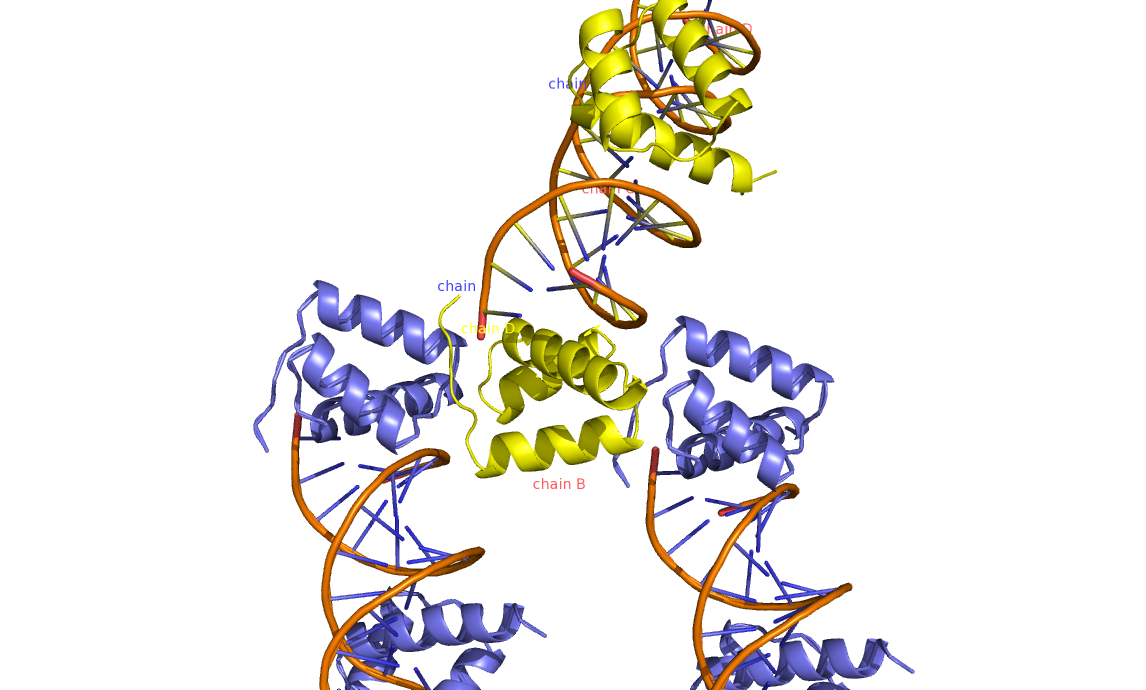
Рис 7. Молекулы ДНК и связанных с ними белками соседних ячеек структуры 3HDD (Желтым обозначена исходная структура, фиолетовым - структуры соседних ячеек).
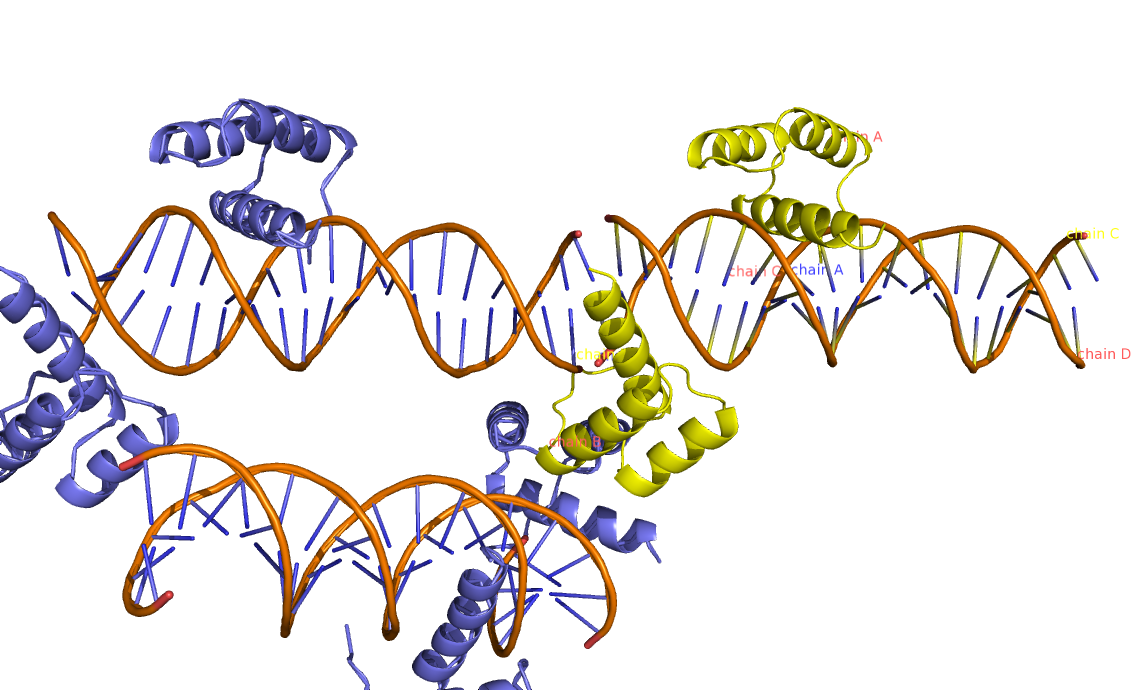
Рис 8. Молекулы ДНК и связанных с ними белками соседних ячеек структуры 3HDD (Желтым обозначена исходная структура, фиолетовым - структуры соседних ячеек) .
Далее были исследованы контакты исходной структуры с соседней ячейкой. Были найдены водородные связи между положительно заряженными остатками и отрицательно заряженным сахарофосфатным остовом ДНК. На рисунках ниже представлены примеры водородных связей, полученные командами:
create connect1, byres (/3HDD//B within 3.5 of sym*)
create connect2, byres (sym* within 3.5 of /3HDD//B)
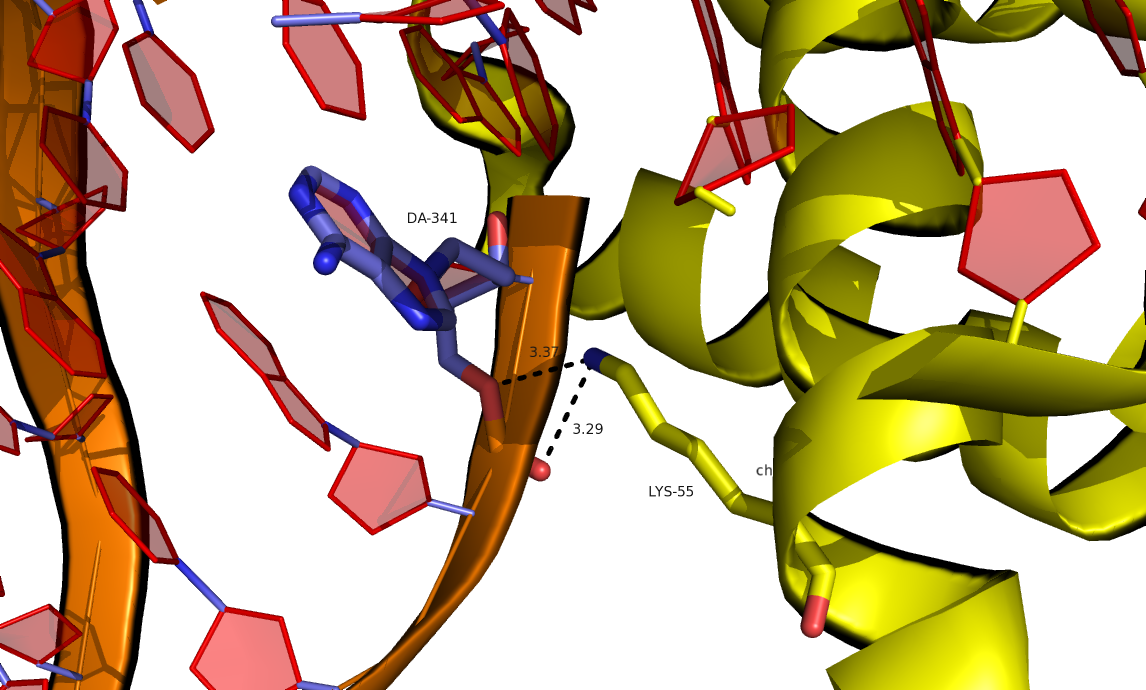
Рис 9. Водородные связи между лизином цепи В исходной структуры и ДНК соседней ячейки.
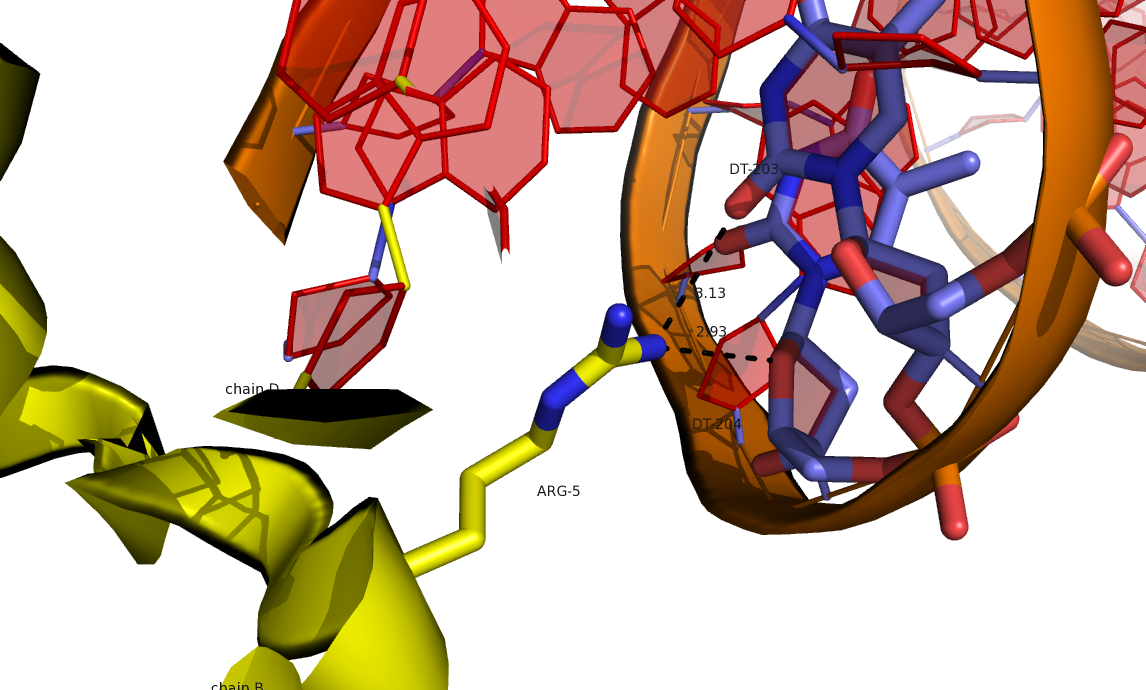
Рис 10.Водородные связи между аргинином цепи В исходной структуры и ДНК соседней ячейки.
Таким образом, странность расположения цепи В белка "на краю" ДНК объясняется тем, что она образует водородные связи с молекулами ДНК соседних ячеек.
Ссылка на проект в PyMol:pr2_3hdd.pse