Исследование ДНК-белковых взаимодействий
Краткое описание структуры в файле 1TRO.pdb
В файле приведены координаты атомов следующих молекул (молекула/цепи):
- TRYPTOPHAN(TRP) REPRESSOR/A, C, E, G
- DNA/I, J, K, L
Для исследования были выбраны цепь А белка триптофанового репрессора и цепи I, J, представляющие молекулу ДНК со следующей последовательностью:
цепь I [1] 5'-TGTACTAGTTAACTAGTAC-3' [19] |||||||||||||||||| цепь J [1] 3'-CATGATCAATTGATCATGT-5' [19],
- Функции белка, структура которого представлена в файле 1TRO.pdb
В файле PDB имеется ссылка на документ UniProt, его accession number P0A881
Белок представлен в виде димера.
Функции: Белок TRPR_ECOLI является апорепрессором, т.е. соединяясь с L-триптофаном, подвергается аллостерической трансформации, которая позволяет ему соединяться с оператором триптофанового оперона (5'-ACTAGT-'3) и ингибировать транскрипцию генов. - Исследование структуры ДНК
С помощью программ find_pair и analyze были получены значения торсионных углов.
Выходной файл: 1tro_aij_old.out
На основе результатов можно сказать, что данная структура ДНК относится к B-форме. Средние значения торсионных углов для внутренних нуклеотидов предвставлены в файле Excel: torsion.xls
Самым "кривым" нуклеотидом оказался 13 цитозин цепочки J. Его суммарные значения торсионных углов, наиболее отклонялись от средних, что можно увидеть в excel на листе "2 метод" из графика. Все отклонившиеся нуклеотиды находятся в близи с белком, что приводит к их не естественным значениям торсионных углов. - Исследование природы ДНК-белковых контактов
Множества, определенные в скрипте my_dna.def:
- polar_atoms - полярные атомы (кислород и азот)
- unpolar_atoms - неполярные атомы (углерод, фосфор и сера)
- polar_dna - атомы ДНК, участвующие в полярных контактах с атомами белка
- unpolar_dna - атомы ДНК, участвующие в неполярных контактах с атомами белка
Контакты атомов белка с остатками: Полярные Неполярные Всего 2'-дезоксирибозы 0 12 12 фосфорной кислоты 15 12 27 азотистых оснований
со стороны большой бороздки2 4 6 азотистых оснований
со стороны малой бороздки0 0 0
Больше всего образуется полярных связей. В основном они в большой бороздке. Также белок "предпочитает" остов для связи нежели основания. - Получение популярной схемы ДНК-белковых контактов с помощью nucplot
Для получения схемы выполнил команду nucplot 1tro_ofs_old.pdb. Конвертировав PS файл, получил популярные схемы ДНК-белковых контактов.
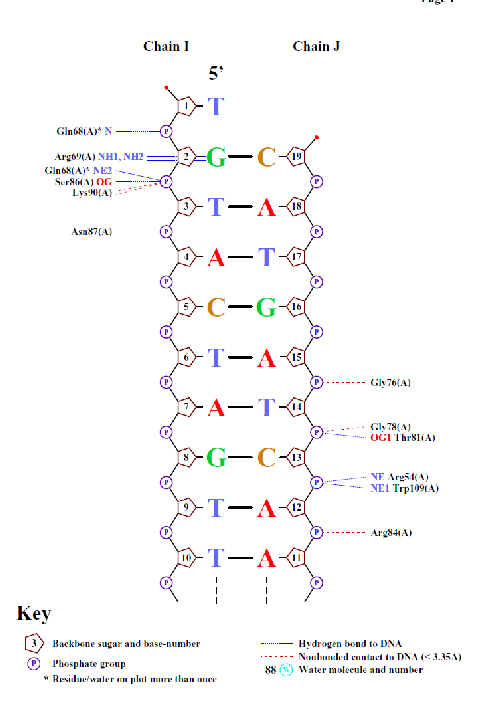
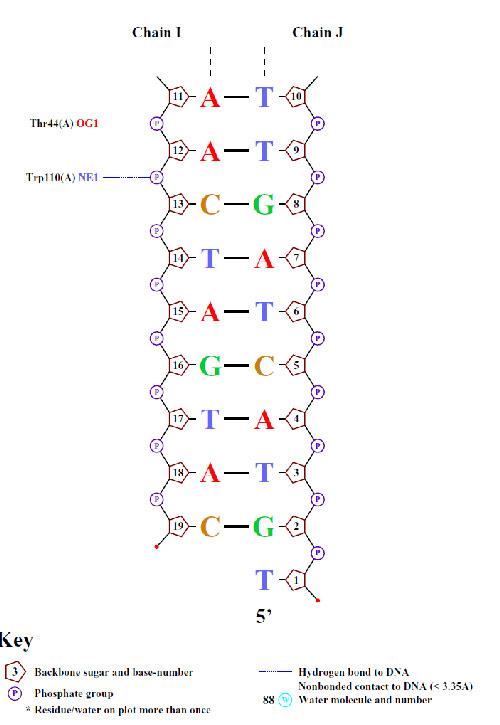
Расчитанный наиболее "кривой" нуклеотид как раз-таки предсказан схемой. Также здесь имеются те нуклеотиды, для которых торсионные углы также отличаются от среднего значения. - Возможный распознающий контакт
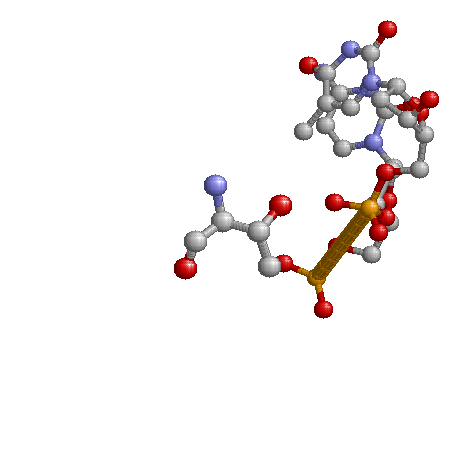
Взаимодействие 2'-дезоксицитозина 13 цепи J и треонина 81 цепи A, является хорошим возможным распознающим контакт. В связи с
наличием водородной связи и рассхождения торсионных углов цитозина 13 цепи J от среднего.
- Характеристика ДНК-связывающего домена P0A881: http://pfam.sanger.ac.uk//protein/P06536
.jpg)
Полное название ДНК-связывающего домена: триптофановый рецептор.
Краткая аннотация домена в InterPro:
InterPro entry IPR000831
The Trp repressor (TrpR) binds to at least five operators in the Escherichia coli genome, repressing gene expression. The operators at which it binds vary considerably in DNA sequence and location within the promoter; when bound to the Trp operon it recognises the sequence 5'-ACTAGT-3' and acts to prevent the initiation of transcription. The TrpR controls the trpEDCBA (trpO) operon and the genes for trpR, aroH, mtr and aroL, which are involved in the biosynthesis and uptake of the amino acid tryptophan PUBMED:12475235. The repressor binds to the operators only in the presence of L-tryptophan, thereby controlling the intracellular level of its effector; the complex also regulates Trp repressor biosynthesis by binding to its own regulatory region. TrpR acts as a dimer that is composed of identical 6-helical subunits, where four of the helices form the core of the protein and intertwine with the corresponding helices from the other subunit.