Исследование ДНК-белковых взаимодействий в структуре комплекса бактериофага лямбда cl-NTD в комплексе с сигма- регионом 4 грамотрицательной палочковидной бактерии рода Thermus
Краткое описание структуры в файле 1rio.pdb
В файле приведены координаты атомов следующих молекул:1.CI белок лямбды бактериофага
2. Cигма фактор РНК полимеразы (H)
3. Репрессор белка CI (цепи А В)
4. N-конец домена
5. 2 молекулы ДНК (T, U)
Организм: Thermus aquaticus (гаммотрицательная палочковидная бактерия )
Белок состоит из 3-х цепей ( A, B, H). Для работы мы будем использовать цепь Н.
ДНК состоит из 2-х цепей (U, T).
Для исследования были выбраны H-цепь белка и цепи T, U, представляющие ДНК со следующей последовательностью:
цепь T [1] CGGTATCACCGCCAGTGCTTGACATGG [27]
|||||||||||||||||||||||||||
цепь U [27] GCCATAGTGGCGGTCACGAACTGTACC [1]
1 и 27 - номера первого и последнего нуклеотида.
Функции белка, структура которого представлена в файле 1rio.pdb
В файле pdb представлен сигма фактор Рнк полимеразы.Его accession number Q9EZJ8. Вы сами можете обо всём узнать на сайте. Фрагмент, который нам интересен небольшой, расположен в белке на 366-438 позициях.теперь подробно изучим его функции.Для связывания с промоторными областями ДНК, основной фермент нуждается в еще одной субъединице - сигма. Сигма-фактор значительно снижает сродство РНК-полимеразы к неспецифичным областям ДНК, и в то же время повышает ее чувствительность к определенным промоторам, в зависимости от своей структуры. С его помощью транскрипция начинается с нужного участка ДНК.
Исследование структуры ДНК
С помощью программ find_pair и analyze определяем тип формы ДНК. Определияем средние значения торсионных углов для внутренних нуклеотидов (для всех, кроме краевых).Для этого нам понадобится файл. Импортируем значения торисионных углов в Excel. Я составил 3 таблички:Первая таблица из файла 1RIO_NUC_old.out.
Во второй таблице все углы пересчитаны в неотрицательные значения, что позволяет высчитывать средние углы.
В третьей таблице находятся величины, характеризующие модуль отклонения величины угла от соответствующего среднего значения. Самый "кривой" нуклеотид со значениями торсионных углов, наиболее отклоняющимися от средних - С21 с цепи U (со средним отклонением, равным 572). Я думаю, что торсионные углы не так сильно зависят от конформации структуры, ведь для этого существуют различные (водородные, стекинг) взаимодействия. Связывание ДНК с белком цепи Н приводит к деформации ДНК для образования наиболее энергенически выгодной структуры.
Исследование природы ДНК-белковых контактов
Будем считать полярными атомы кислорода и азота, а неполярными - атомы углерода, фосфора и серы. Назовем полярным контактом ситуацию, в которой расстояние между полярным атомом белка и полярным атомом ДНК меньше 3.5A. Аналогично, неполярным контактом будем считать пару неполярных атомов на расстоянии меньше 4.5A. Скрипт..Таблица. Контакты разного типа в комплексе 1rio.pdb
| Контакты атомов белка с | Полярные | Неполярные | Всего |
| остатками 2'-дезоксирибозы | 0 | 22 | 22 |
| остатками фосфорной кислоты | 24 | 11 | 35 |
| остатками азотистых оснований со стороны большой бороздки | 16 | 35 | 51 |
| остатками азотистых оснований со стороны малой бороздки | 1 | 1 | 2 |
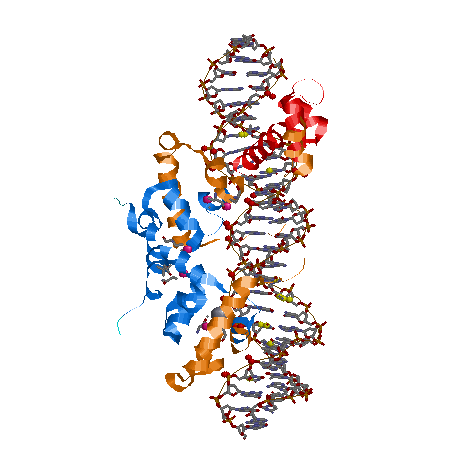
Сделаем вывод. Во-первых полярных молекул на много меньше, чем неполярных.Белок взаимодействует с фосфатами намного больше, чем с сахаром.Это обусловлено тем, что фосфаты находятся снаружи спирали. Что касается большой и малой бороздки, то контактов больше с ао большой бороздки.Это связано с глубиной б.бороздки. На� картинке представлены различные контакты. Желтым указаны атомы ДНК, участвующие в полярных контактах с атомами белка.
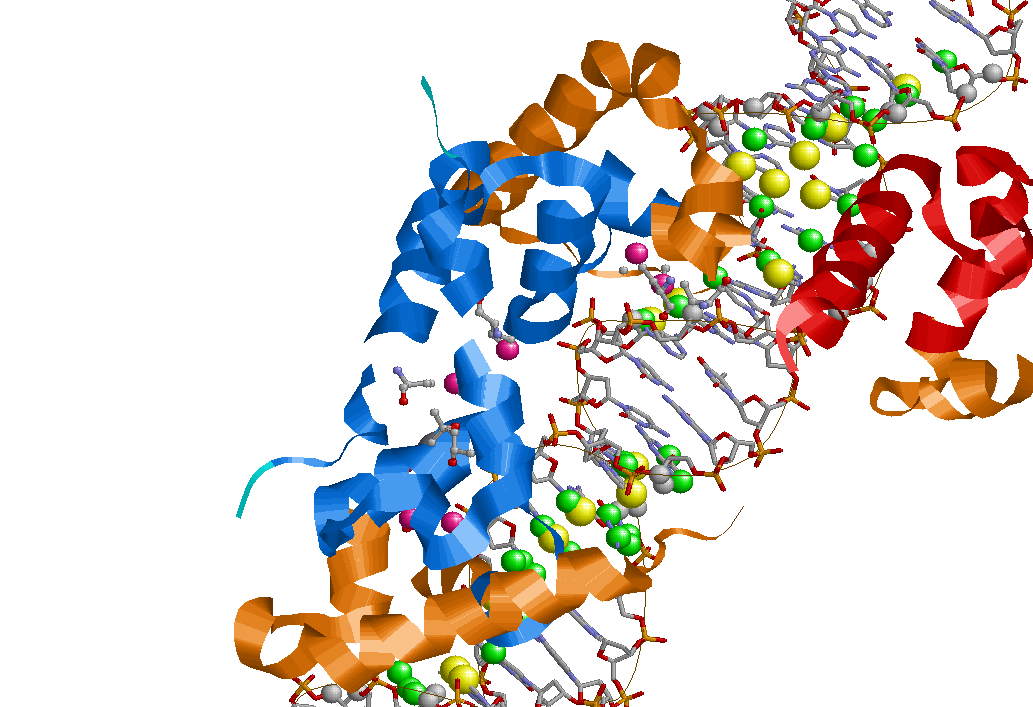
Получение популярной схемы ДНК-белковых контактов с помощью nucplot
Для получения популярной схемы ДНК-белковых контактов была выполнена следующая команда: nucplot 1RIO_old.pdb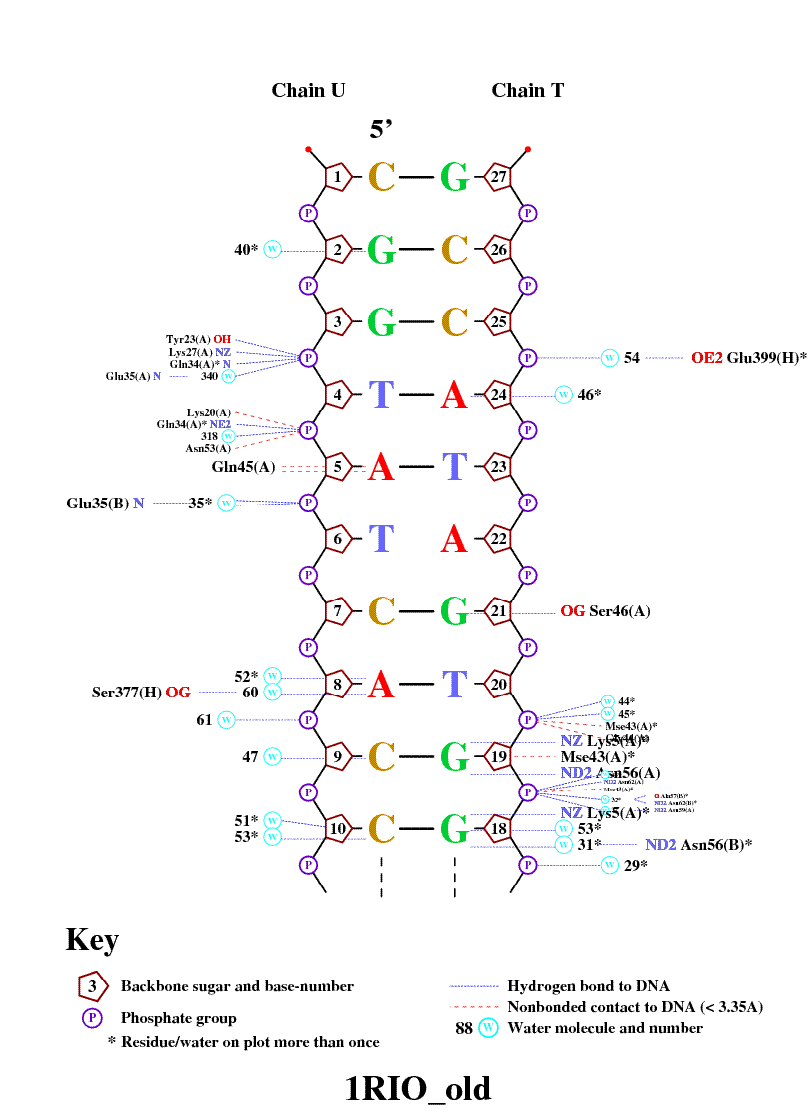
Как только я получил изображение с различными контактами ДНК с белком, меня немного смутило, что контакты немного не совпадают с теми, что я делал в предыдущих упражнениях. Может быть это связано с тем, что программа nucplot определяет взаиможействия более сложным способом.
Возможный распознающий контакт
Распознающим контактом является контакт A5:U - Gln45:A, так как в нем участвуют сразу 2 водородные связи, а в остальных случаях учавствует одна водородная связь.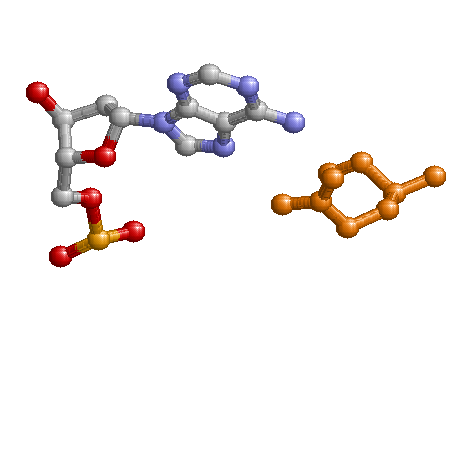
Характеристика ДНК-связывающего домена Q9EZJ8
Cначала перейдём по ссылке.
Полное название ДНК-связывающего домена: RNA polymerase sigma-70 region 4
1)The bacterial core RNA polymerase complex, which consists of five subunits, is sufficient for transcription elongation and termination but is unable to initiate transcription. Transcription initiation from promoter elements requires a sixth, dissociable subunit called a sigma factor, which reversibly associates with the core RNA polymerase complex to form a holoenzyme [1]. RNA polymerase recruits alternative sigma factors as a means of switching on specific regulons. Most bacteria express a multiplicity of sigma factors. Two of these factors, sigma-70 (gene rpoD), generally known as the major or primary sigma factor, and sigma-54 (gene rpoN or ntrA) direct the transcription of a wide variety of genes. The other sigma factors, known as alternative sigma factors, are required for the transcription of specific subsets of genes. With regard to sequence similarity, sigma factors can be grouped into two classes, the sigma-54 and sigma-70 families. Sequence alignments of the sigma70 family members reveal four conserved regions that can be further divided into subregions eg. sub-region 2.2, which may be involved in the binding of the sigma factor to the core RNA polymerase; and sub-region 4.2, which seems to harbor a DNA-binding 'helix-turn-helix' motif involved in binding the conserved -35 region of promoters recognised by the major sigma factors [2, 3]. Region 4 of sigma-70 like sigma-factors is involved in binding to the -35 promoter element via a helix-turn-helix motif [4]. Due to the way Pfam works, the threshold has been set artificially high to prevent overlaps with other helix-turn-helix families. Therefore there are many false negatives.
2)Наиболее изученной из бактериальных ферментов является РНК- полимераза E.coli. Она осуществляет транскрипцию всех бактериальных генов. Фермент состоит из пяти субъединиц: бета'- (молекулярная масса 165 кДа), бета- (155 кДа), двух альфа- (35 кДа каждая) и сигма- (чаще всего 70 кДа (сигма70)). Комплекс из четырех субъединиц бета-бета'-альфа-альфа, часто обозначаемый буквой Е (enzyme), образует так называемый минимальный (кор-) фермент E.coli , который способен осуществлять все основные этапы транскрипции, за исключением правильной инициации. Для инициации транскрипции требуется присутствие определенной регуляторной сигма-субъединицы, необходимой для распознавания РНК-полимеразой промоторов бактериальных генов, определяющей специфичность взаимодействия РНК-полимеразы с промоторами и, возможно, последующую изомеризацию комплекса РНК-полимераза-промотор, необходимую для начала синтеза РНК. Полный фермент, включающий сигма70-субъединицу, часто называют холоферментом и обозначают Есигма70. РНК-полимераза Есигма70 способна транскрибировать большинство (но не все) генов E.coli. В частности, для транскрипции генов теплового шока , оперонов gln или nif требуется включение в состав полного фермента другой регуляторной субъединицы - сигма54 (молекулярная масса 54 кДа) вместо сигма70 с образованием фермента Eсигма54 . Описано до десяти различных сигма-факторов, объединение которых с минимальным ферментом дает возможность образующимся холоферментам узнавать разные промоторы. Все четыре субъединицы кор-фермента обеспечивают контакт РНК-полимеразы с промоторами. При этом бета'-субъединица участвует в связывании фермента с ДНК, бета-субъединица образует каталитический активный центр, а альфа- субъединицы обеспечивают правильное взаимодействие фермента с промоторами. Данные такого рода получают с использованием ферментов, у которых под действием мутаций изменены конкретные субъединицы, и если, например, мутация в гене альфа-субъединицы нарушает связывание РНК-полимеразы с ДНК, делаются соответствующие выводы. Однако любая мутантная субъединица в составе олигомерного фермента может изменять его конформацию и придавать ферменту самые неожиданные свойства. Более прямым методом определения мест контакта макромолекул при белок-белковых и белково-нуклеиновых взаимодействиях является метод поперечных сшивок с использованием бифункциональных химических агентов. Такие химические соединения образуют ковалентные связи (поперечные сшивки) между близкорасположенными реакционноспособными группами. Наличие контакта между макромолекулами нельзя однозначно интерпретировать в пользу его функциональной значимости.