Учебный сайт Валяевой Анны
A- и В- формы ДНК. Структура РНК
Задание 1
С помощью инструментов пакета 3DNA, работающего под LINUX, и программы fiber были построены модели структур A-, B- и Z-формы ДНК. В качестве повторяющегося элемента был выбран олигонуклеотид GATC, колчество его повторений - 5 (для Z-формы 6). В результате были получены 3 pdb-файла: gatc-a.pdb, gatc-b.pdb, gatc-z.pdb.
Задание 2
На рисунке 1 представлено изображение структуры фрагмента ДНК A-формы, созданное в Jmol. Файл pdb был взят из предыдущего задания.
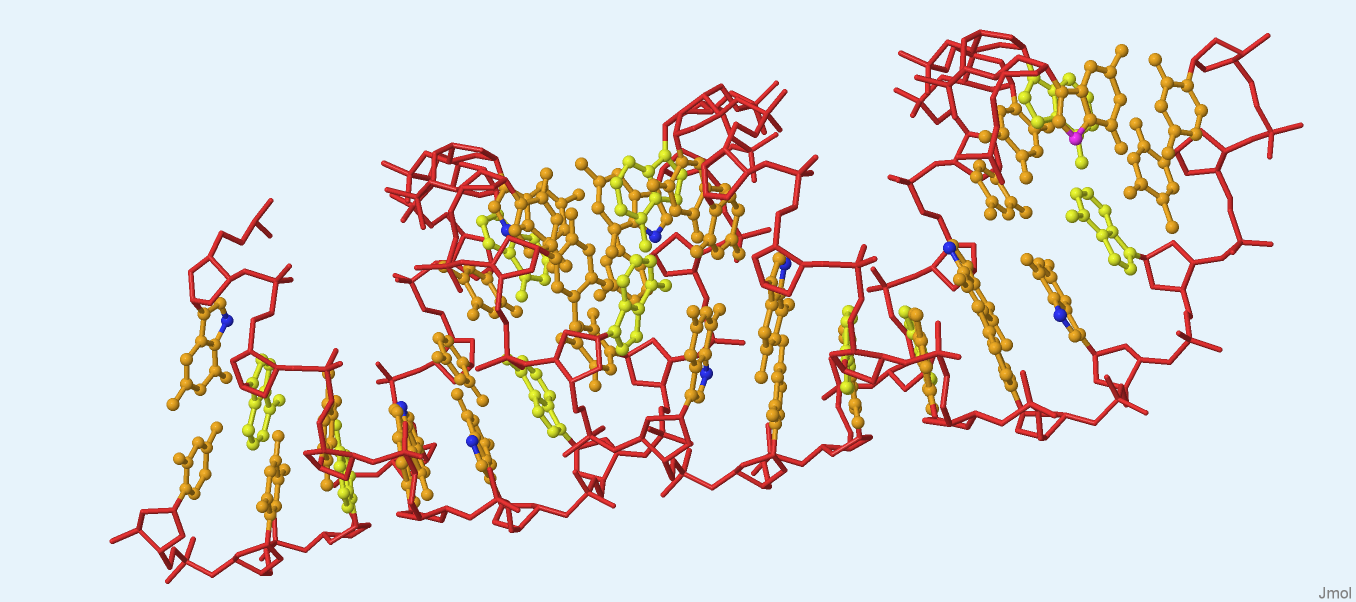
Рис. 1. Структура фрагмента ДНК A-формы. Красным цветом раскрашен сахарофосфатный остов, оранжевым - азотистые основания, желтым - аденины, синим - атомы N7 гуанинов, розовым - атом N7 первого в последовательности гуанина. Изображение получено с помощью программы Jmol.
Далее были получены изображения только ДНК и только РНК (рис. 2, 3) из файлов 1by4.pdb1 и 2dxi.pdb1, соответственно. Разрывов обнаружено не было. Координаты атомов ДНК и РНК записаны в файлах dna.pdb и rna.pdb, соответственно.
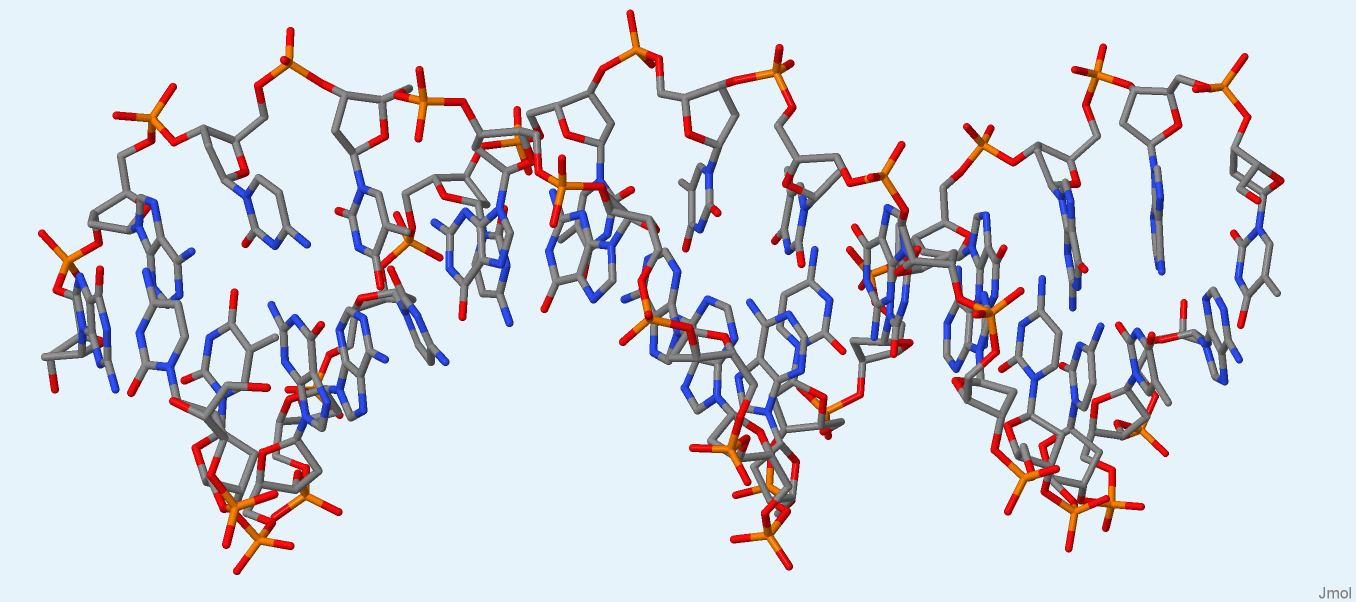
Рис. 2. ДНК из структуры 1by4. Изображение получено с помощью программы Jmol.
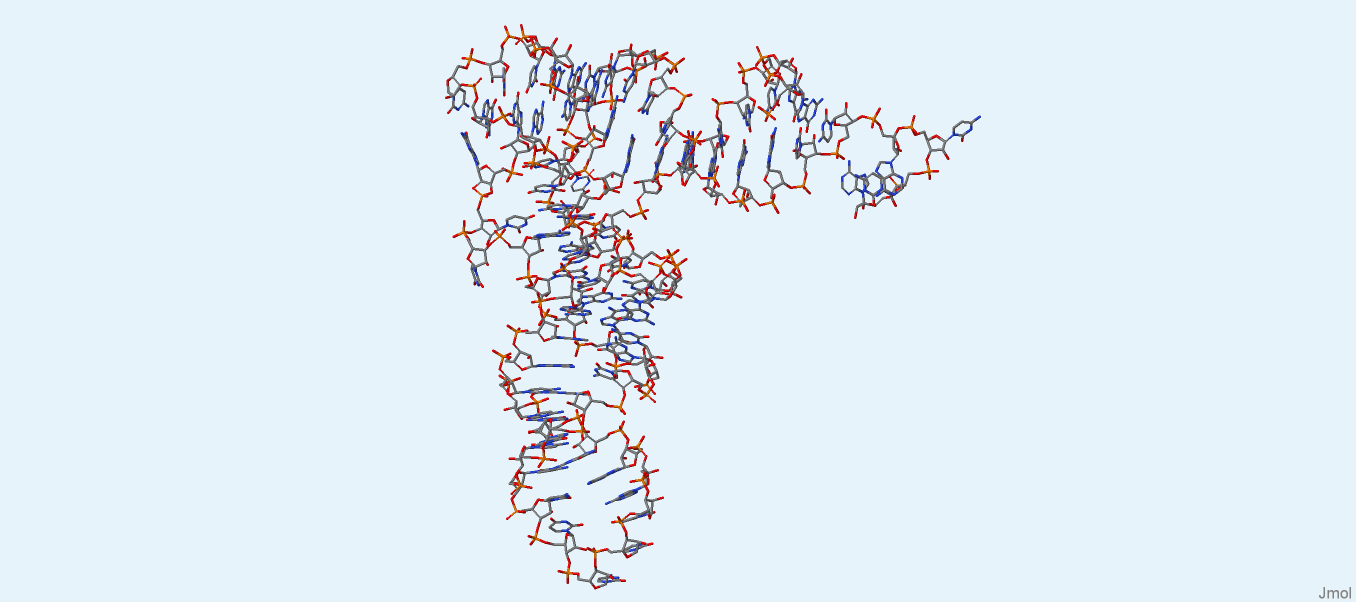
Рис. 3. РНК из структуры 2dxi. Изображение получено с помощью программы Jmol.
Задание 3
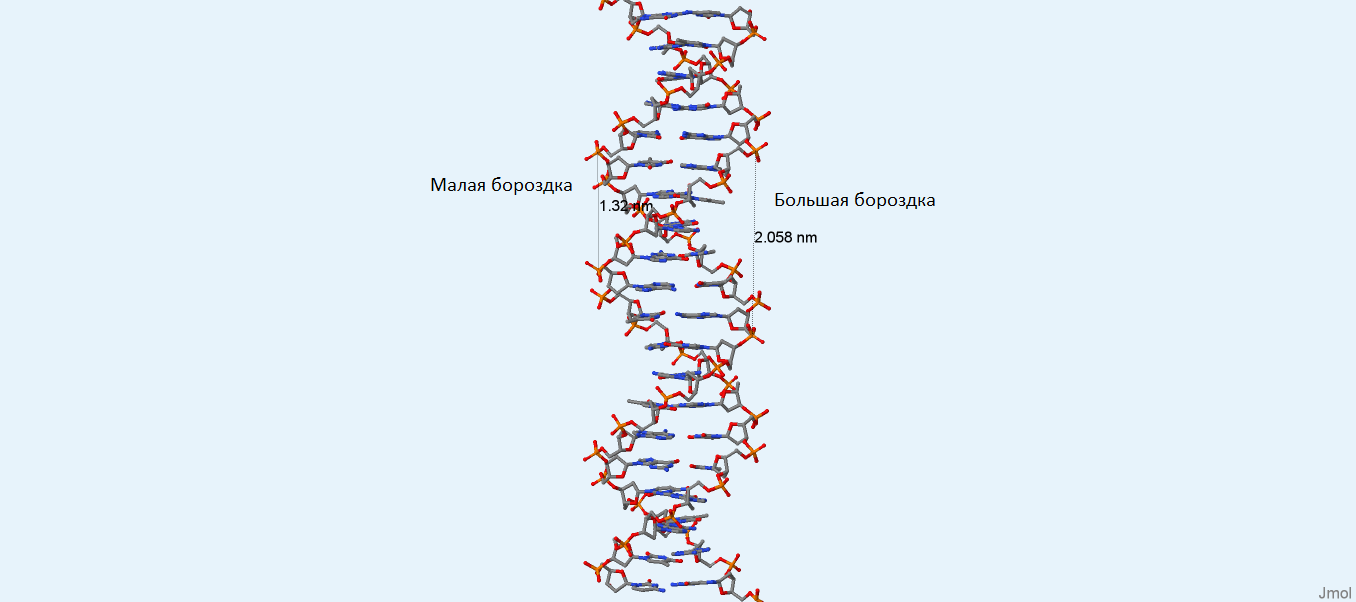
Рис. 4. Большая и малая бороздки B-формы ДНК. Изображение получено с помощью программы Jmol.
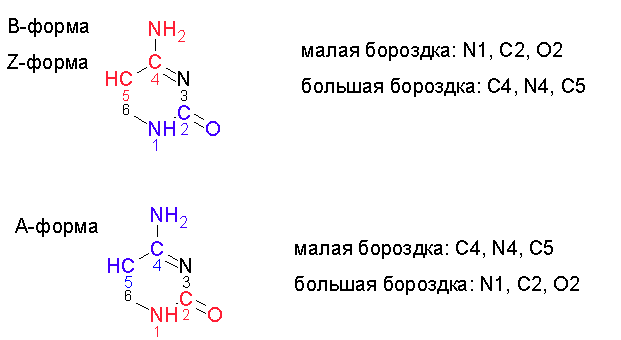
Рис. 5. Ориентация атомов цитозина относительно большой и малой бороздок. Красным цветом раскрашены атомы, смотрящие в сторону большой бороздки, синим - в сторону малой. Изображение получено с помощью программы ChemSketch.
В таблице 1 представлено сравнение некоторых параметров структуры A-, B- и Z-форм ДНК. Для ширины большой и малой бороздок указаны номера нуклеотидов, меджу фосфатами которых измрялось расстояние.
Таблица 1. Сравнение параметров форм ДНК.
| A-форма | B-форма | Z-форма | |
| Тип спирали | правая | правая | левая |
| Шаг спирали (A) | 28.03 | 33.75 | 43.5 |
| Число оснований на виток | 11 | 10 | 12 |
| Ширина большой бороздки | 16.81 (T27:B-C16:A) | 17.91 (A6:A-T31:B) | 18.3 (C32:B-C6:A) |
| Ширина малой бороздки | 7.98 (C8:A-T27:B) | 11.69 (T31:B-A14:A) | 7.2 (G13:A-G31:B) |
Далее были измерены торсионные углы C32 в A- и B-формах ДНК в Jmol. Сравнение полученных результатов с данными из презентации представлено в табл. 2.
Таблица 2. Торсионные углы C32.
| α | β | γ | δ | ε | ζ | χ | |
| A-форма из презентации | 62 | 173 | 52 | 88/3 | 178 | -50 | -160 |
| A-форма | -51.7 | 174.8 | 41.7 | 79.1/-159.3 | -147.8 | -75.1 | -94.9 |
| B-форма из презентации | 63 | 171 | 54 | 123/131 | 155 | -90 | -117 |
| B-форма | -29.9 | 136.3 | 31.1 | 143.3/-100.7 | -140.8 | -160.5 | -97.9 |
Задание 4
Для получения файла со значениями торсионных углов в заданных структурах ДНК и тРНК были использованы следующие команды:
remediator --old dna.pdb > dna_old.pdb find_pair dna_old.pdb stdout | analyze find_pair -t rna.pdb stdout | analyze
Далее полученные данные были обработаны в MS Excel (полученный файл), средние значения торсионных углов представлены в таблице 3.
Таблица 3. Средние значения торсионных углов в ДНК 1by4 и тРНК 2dxi.
| α | β | γ | δ | ε | ζ | χ | |
| 1by4 | -2,2 | 12,5 | 9 | 144 | -92,2 | -86,7 | -98,5 |
| 2dxi | -47,8 | 71,5 | 48,3 | 85,5 | -150,1 | -71,6 | -133,8 |
Исходя из полученных результатов, можно говорить что наиболее "деформированный" нуклеотид в заданной ДНК - это цитозин 13 из второй цепи. А структура заданной тРНК больше всего напоминает А-форму ДНК.
Водородные связи в тРНК
Акцепторный стебель (0.010) C:.501_:[..G]G-----C[..C]:.572_:C (0.006) (0.014) C:.502_:[..G]G-*---U[..U]:.571_:C (0.008) (0.008) C:.503_:[..C]C-----G[..G]:.570_:C (0.010) (0.004) C:.504_:[..C]C-----G[..G]:.569_:C (0.008) (0.010) C:.505_:[..C]C-----G[..G]:.568_:C (0.010) (0.006) C:.506_:[..C]C-----G[..G]:.567_:C (0.007) (0.008) C:.507_:[..A]Ax----U[..U]:.566_:C (0.006)
Т-стебель (0.007) C:.549_:[..G]G-----C[..C]:.565_:C (0.004) (0.004) C:.550_:[..G]G-----C[..C]:.564_:C (0.007) (0.006) C:.551_:[..G]G-----C[..C]:.563_:C (0.003) (0.004) C:.552_:[..G]G-----C[..C]:.562_:C (0.005) (0.004) C:.553_:[..G]G----xC[..C]:.561_:C (0.004)
Акцепторный стебель (0.007) C:.538_:[..A]A-*---C[..C]:.532_:C (0.004) (0.005) C:.539_:[..G]G-----C[..C]:.531_:C (0.004) (0.010) C:.540_:[..G]G-----C[..C]:.530_:C (0.007) (0.005) C:.541_:[..C]C-----G[..G]:.529_:C (0.004) (0.003) C:.542_:[..C]C-----G[..G]:.528_:C (0.007) (0.009) C:.543_:[..G]G-----C[..C]:.527_:C (0.006) (0.006) C:.544_:[..A]Ax*---G[..G]:.526_:C (0.008)
D-стебель (0.009) C:.510_:[..G]G-----C[..C]:.525_:C (0.007) (0.007) C:.511_:[..U]U-----A[..A]:.524_:C (0.009) (0.008) C:.512_:[..C]C-----G[..G]:.523_:C (0.013) (0.004) C:.513_:[..U]U-*--xG[..G]:.522_:C (0.009)
Структуру стеблей поддерживают 19 канонических пар оснований и 4 неканонические пары. В поддержании третичной структуры участвуют пары, не задействованные в образовании стеблей: 554U-A558, 555U-G518, 519G-C556, 514A-U508, 520U-C548.
Для следующих пар последовательных пар азотистых оснований были определены наибольшие значения площадей перекрывания.
step i1-i2 i1-j2 j1-i2 j1-j2 sum
2 GC/GU 7.07( 4.32) 0.00( 0.00) 0.00( 0.00) 6.67( 4.16) 13.74( 8.48)
13 UU/GA 5.74( 2.64) 0.00( 0.00) 0.00( 0.00) 5.06( 2.34) 10.80( 4.98)
16 AG/CC 4.41( 2.45) 0.00( 0.00) 0.00( 0.00) 5.62( 2.75) 10.03( 5.20)
18 GC/GC 6.88( 3.75) 0.00( 0.00) 0.00( 0.00) 6.56( 3.44) 13.44( 7.19)
Изображения стекинг-взаимодействий данных пар оснований, полученные с помощью команд, приведенных ниже, представлены на рисунках 6-9.
ex_str -2 stacking.pdb step2.pdb stack2img -cdolt step2.pdb step2.ps convert step2.ps step2.png, где 2 - номер одного из шагов (step)
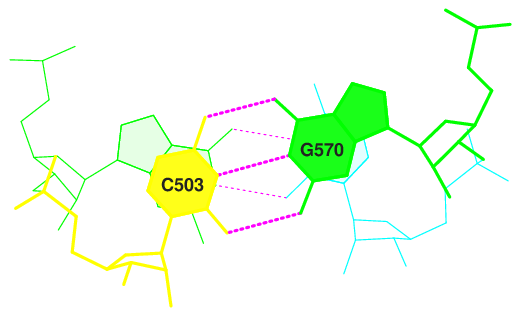
Рис. 6. Стекинг-взаимодействия в структуре тРНК 2dxi. Шаг 2.
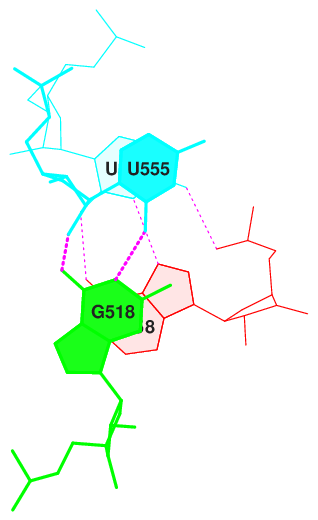
Рис. 7. Стекинг-взаимодействия в структуре тРНК 2dxi. Шаг 13.
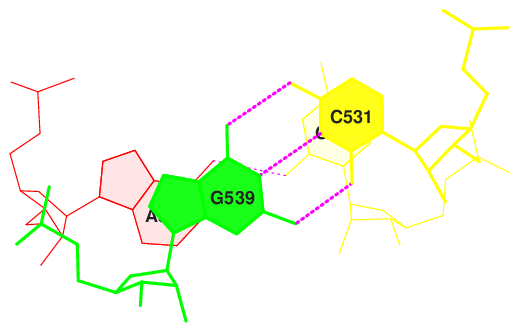
Рис. 8. Стекинг-взаимодействия в структуре тРНК 2dxi. Шаг 16.
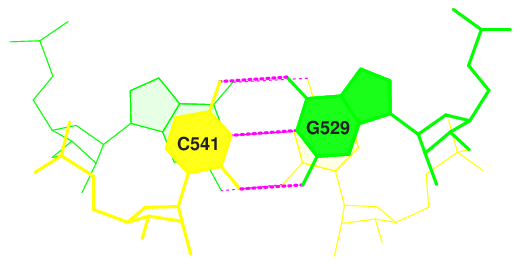
Рис. 9. Стекинг-взаимодействия в структуре тРНК 2dxi. Шаг 18.
Дата последнего обновления: 26.10.14
©Валяева Анна