Восстановление кристалла из PDB файла
|
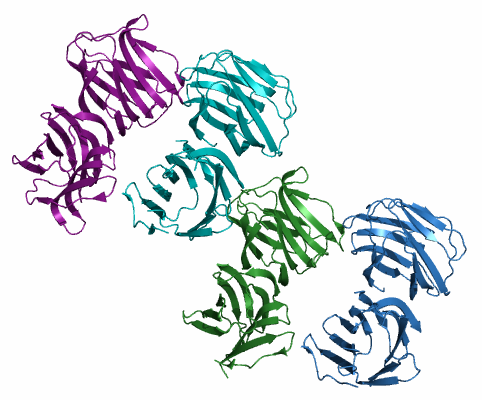
| Рис. 1. Структура молекул белка из одной кристаллической ячейки. Разные гомодимеры покрашены разным цветом. |
Структура кристалла галектина-1
Продолжим исследование структуры крысиного галектина-1 (PDB код 3m2m). В поле CRYST1 PDB-файла содержится следующая запись:
40.100 58.600 121.600 101.10 91.60 110.90 P 1 8
Отсюда характеристики кристалла:
- Длины направляющих векторов кристалла:
- a = 40.1 Å
- b = 58.6 Å
- c = 121.6 Å
- Углы между направляющими векторами кристалла:
- a = 101.1 °
- b = 91.6 °
- c = 110.9 °
- Кристаллографическая группа: Р 1, то есть ячейка примитивная триклинная.
- Число молекул в ячейке: 8
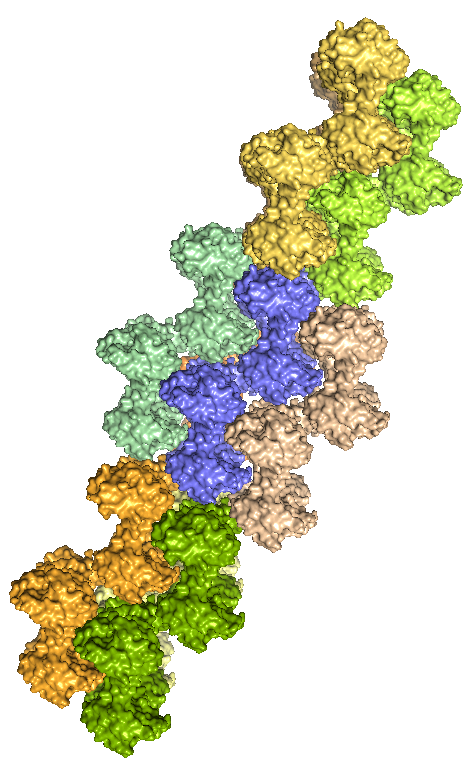
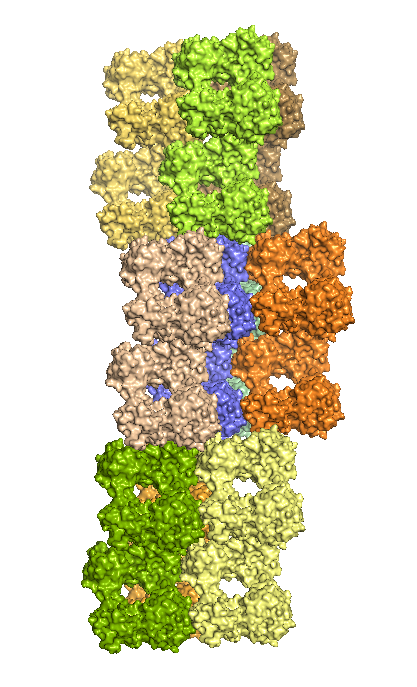
Структуру белка визуализировали в PyMOL. На рисунке 1 приведено изображение содержимого одной кристаллической ячейки, на рисунке 2 - восстановленные командой symexp соседние кристаллические ячейки. Видно, что молекулы из разных ячеек плотно состыкованы, а отдельные субъединицы белка образуют ряды.
|
|
| Рис. 2. Молекулы белка из соседних кристаллических ячеек с разных ракурсов. Каждая ячейка выделена своим цветом. Синим выделена исходная ячейка. | |
Рассмотрим различные контакты между молекулами белка в кристалле.
- Внутри гомодимера. Автором в PDB файле указано, что белок существует в форме димера, и димеры в данном случае образованы парами цепей A и B, C и D, E и F, G и H. Чтобы найти связи между цепями A и B, в PyMOL использовались операторы byres и within и команда distance. Каждая из субъединиц существует в виде β-сэндвича. Мы видим, что между β-листами, принадлежащими разным цепям, но лежащими примерно в одной плоскости, достаточно большое количество водородных связей, которые как бы сдваивают их в один длинный β-лист (см. рис. 3).
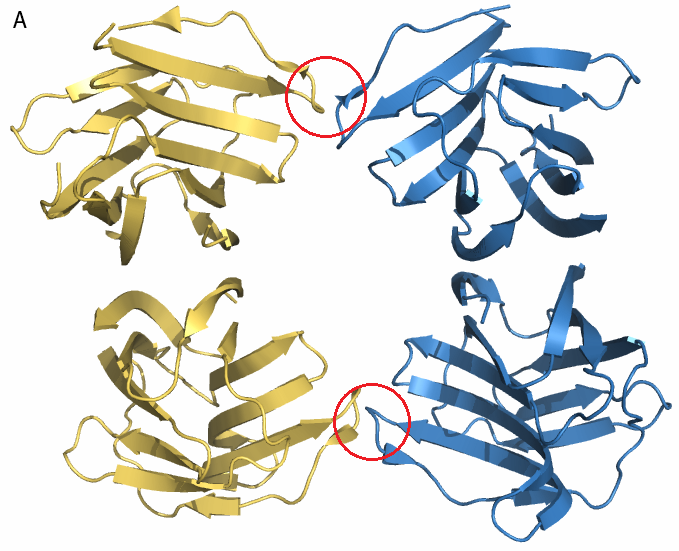
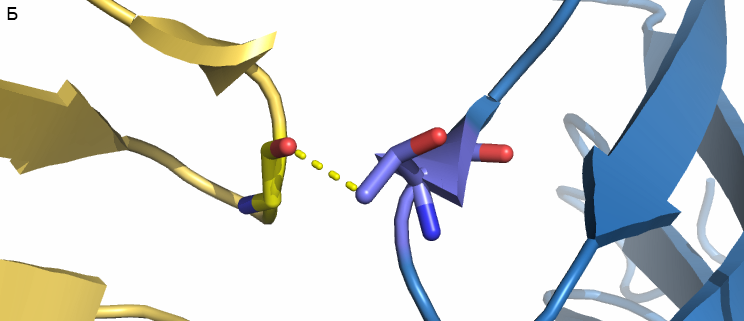
- Внутри ячейки. Каждая ячейка кристалла содержит 4 гомодимера. Как можно увидеть на рисунке 1 димеры могут взаимодействовать "боками" либо "по диагонали". На рисунке 4 приведены гомодимеры, образованные цепями A и B, F и E. Между ними есть только два небольших участка, где могут быть какие-либо связи. На расстоянии меньше 3.5 Å расположены только атом кислорода из Gly66 цепи F и углерод боковой цепи Thr67 цепи B (и два симметричных им атома из цепей А и Е). Диагонально расположенные гомодимеры здесь рассматривать не будем, так как взаимодействия между ними совпадают с взаимодействиями между белками из соседних ячеек.
- Между ячейками. Рассмотрим взаимодействия молекул белка из соседних ячеек. Для удобства сначала рассматривались две ячейки, расположенные одна под другой, а затем ячейки, стыкующиеся боковыми сторонами. В первом случае (см. рис. 5) взаимоействий было найдено не много, только между цепью G одной ячейки и цепью H ругой ячейки. Во втором случае все стыкующиеся боками цепи имеют несколько взаимодействующих атомов. На рисунке 6 приведен пример для двух из таких цепей.
А
|
Б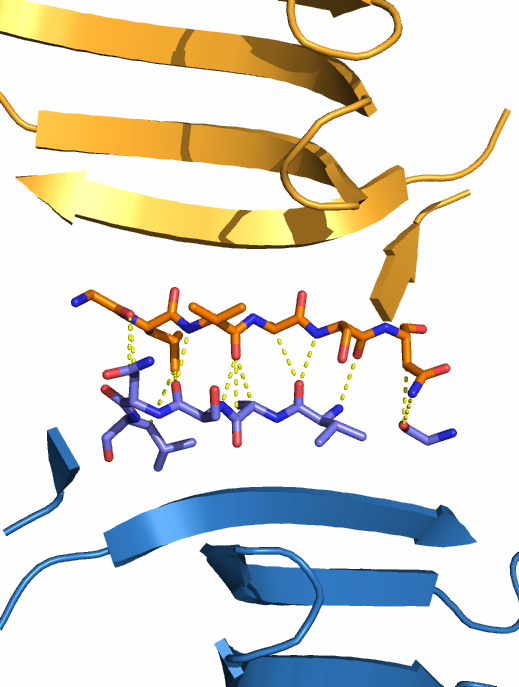
|
В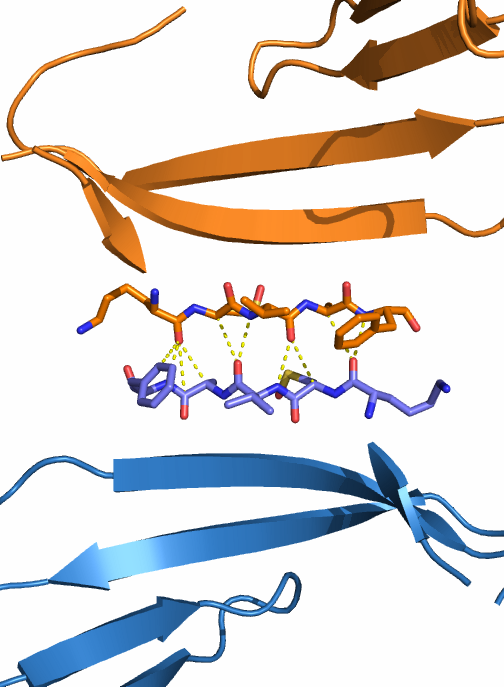
|
| Рис. 3. Водородные связи между субъединицами гомодимера галектина-1. Цепь А показана оранжевым цветом, цепь В - синим. Водородные связи показаны жёлтым пунктиром. А - общий вид на гомодимер; видно, что два ряда водородных связей между субъединицами лежат в плоскостях β-листов. Б и В - половинки гомодимера, "разрезанного" вдоль для удобного отображения водородных связей. | ||
|
|
| Рис. 4. Гомодимеры A+B (синий) и F+E (жёлтый). А - общий вид; красным выделены области контакта. Б - связь между атомами Gly66 цепи F и Thr67 цепи B (показана жёлтым пунктиром); длина связи - 2.7 Å. |
А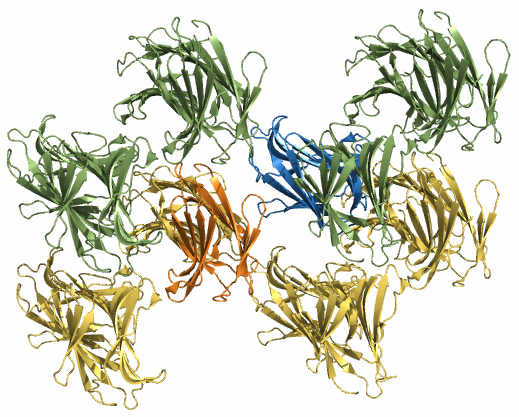
|
Б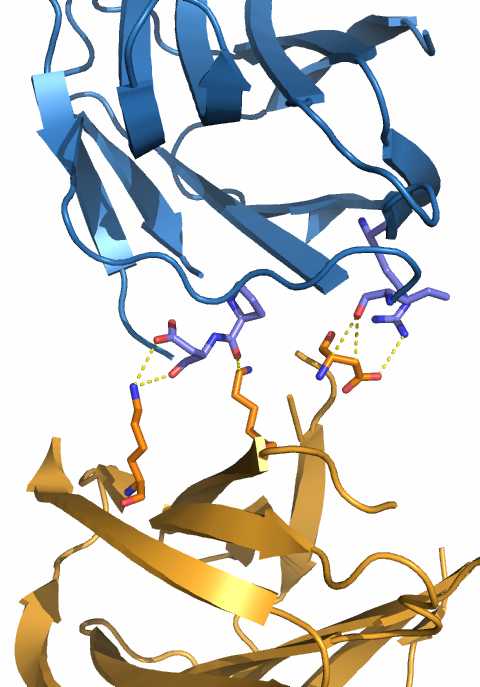
|
| Рис. 5. Молекулы галектина-1 из соседних ячеек (одна показана жёлтым цветом, другая - зелёным). А - общий вид; взаимодействующие цепи выделены цветами - оранжевым цепь G из одной ячейки, синим - цепь H из другой. Б - взаимодействующие остатки (длина связи менее 3.5 Å). | |
| А
|
Б
|
| Рис. 6. Молекулы галектина-1 из соседних ячеек (одна показана жёлтым цветом, другая - зелёным), расположенных в ряд. А - общий вид; взаимодействующие цепи выделены цветами - оранжевым цепь G из одной ячейки, синим - цепь H из другой. Б - взаимодействующие остатки (длина связи менее 3.5 Å). | |
Выводы. Данный белок удалось закристаллизовать, так как отдельные молекулы белка могут образовывать достаточно большое количество связей между собой. Вероятно, контакты между белками, входящими в один гомодимер, по данным PDB файла, отражают их природные взаимодействия.
Структура ДНК-белкового комплекса 3HDD
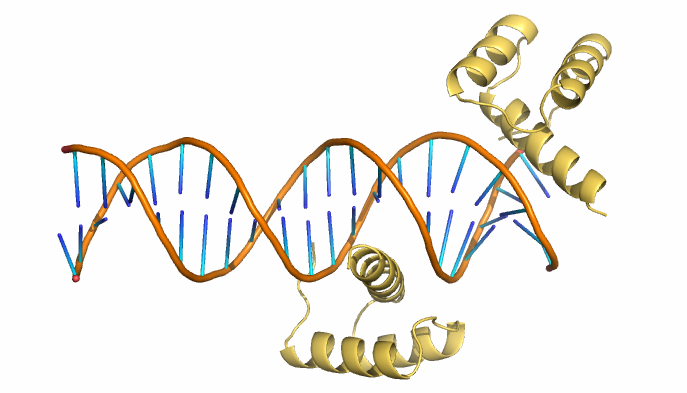
PDB-файл 3HDD содержит структуру комплекса гомеодомена и цепи ДНК из Drosophila melanogaster (см. рис. 7). В нём присутствует "странность" - одна из цепей белка расположена слишком близко к концу цепи ДНК.
|
| Рис. 7. Структура 3HDD. Здесь и далее жёлтым показаны цепи белка, оранжевым - сахарофосфатный остов ДНК, синим - азотистые основания. |
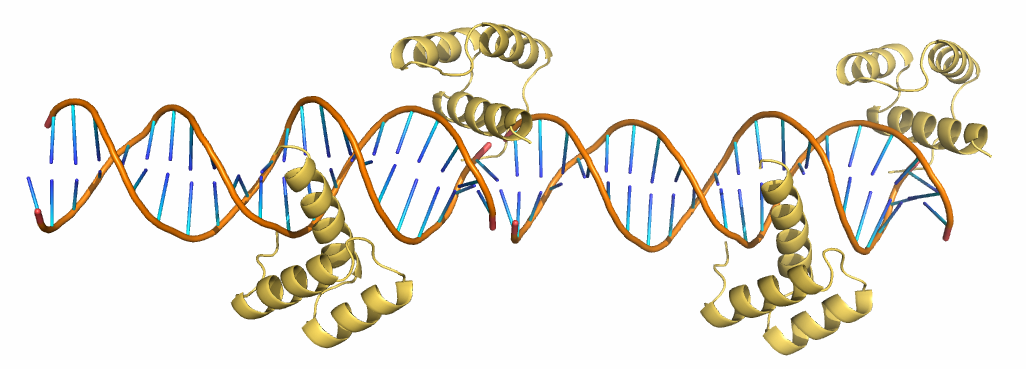
Чтобы понять, почему структура выглядит так, была восстановлена соседняя ячейка (см. рис. 8). Оказалось, что краевые нуклеотиды соседних цепей ДНК образуют Уотсон-Криковскую пару. На самом деле молекула белка расположена не на краю цепи ДНК, а в середине, как и другая.
|
| Рис. 8. Структура 3HDD с восстановленной соседней ячейкой. |
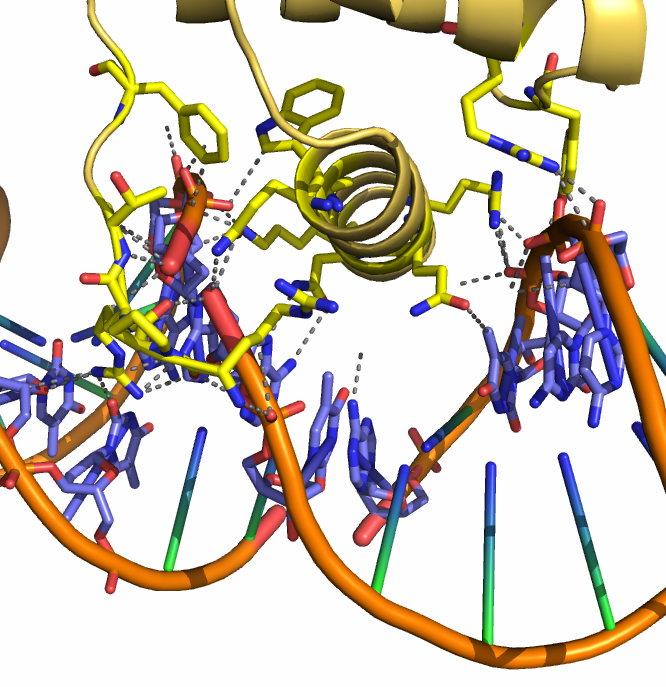
Это подтверждается наличием большого числа контактов между молекулой белка и цепями ДНК из обеих ячеек (см. рис. 9).
|
| Рис. 9. Участок взаимодействия цепи В с ДНК из двух ячеек. Серым пунктиром показаны связи, длина которых меньше 3.5 Å. Взаимодействующие остатки покрашены стандартными цветами химических элементов. |