В данном практикуме я работала со структорой 2AXW из PDB, она соответсвует расшифровке структуры инвазина DraD из уропатогенной Escherichia coli.
Оригинальную публикацию 2006 года, к которой относится эта расшифровка, можно посмотреть или скачать здесь.
Авторы публикации: Robert Je˛drzejczak, Zbigniew
Dauter, Mirosława Dauter,
Rafał Pia˛tek, Beata Zalewska,
Marta Mro´z, Katarzyna Bury,
Bogdan Nowickid and Jo´zef
Kurc.
Результатом исследования, описанного в статье, стало подтверждеение того, что
DraD способен агрегировать, принимая внешнюю
комплементарную донорскую G цепь. Дополнительный C-концевой
участок из 13 аминокислот по счастливой случайности произвел
роль такой донорной цепи, которая позволила молекулам
образовать симметричный димер. Также было подтверждено, что этот белок располагается на конце фибриллы, и кепирует ее кончик (всмысле 'cap').
Кристаллическая структура
конструкции E. coli DraD, обладающая дополнительным
С-концевым удлинением из 13 аминокислот, включая тег His6,
определена при разрешении 1.05 A˚.
Кластер генов dra уропатогенных штаммов Escherichia coli кодирует белки, участвующие в прикреплении бактерий и
вторжении в ткани эукариот-хозяев. Белковые формы
симметрично димеризуются за счет обмена C-концевыми beta-структрурированными участками, которые участвуют в иммуноглобулиноподобных beta-sandwich каждой субъединицы.
Эта структура подтверждает, что
DraD может действовать как акцептор в донорной цепи
механизма комплементации образования волокон, но, в
в отличие от адгезина DraE, его нативная последовательность не имеет
донорскую нить; следовательно, DraD может располагаться только на кончике
волокна. До публикации данной работы структура такого белка еще не была получена.
Отчет о валидации можно посмотреть здесь.
Из отчета можно узнать значения разных метрик валидации структуры, например, значение Rfree фактора равно 0,144.
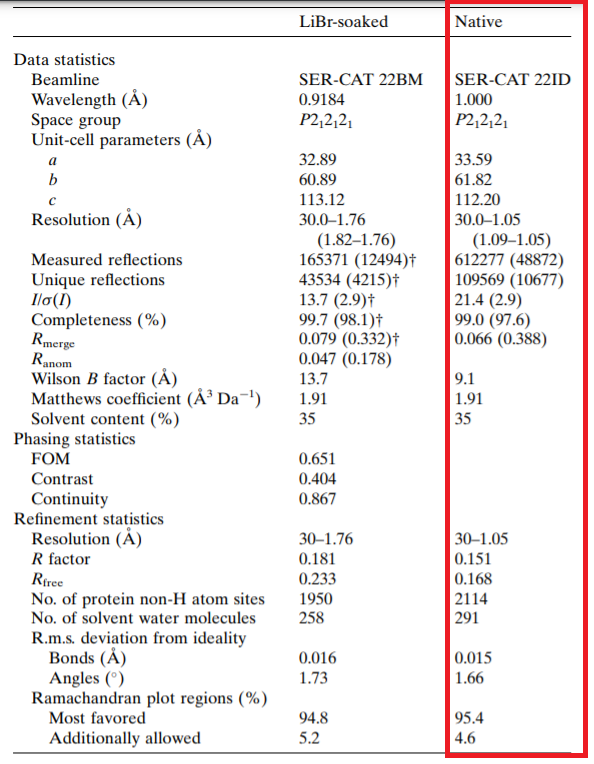
Рис.1. Таблица с дифракционными данными и статистикой уточнения из статьи.
Количество измеренных рефлексов 612277 (48872 для модели с лучшим разрешением (shell)). Полнота данных 99%.
Диапазон разрешений структурных факторов 30–1.05 ангстрем, что достаточно большой диапазон значений. Параметры кристаллографической ячейки
a, b, c, α, β, γ: 33.59Å, 61.82Å, 112.20Å,
90.00◦, 90.00◦, 90.00◦.
Значения индикаторов качества модели в целом. R-фактор равен 0.151, согласно Depositor, или 0.154, согласно с DCC.
R-free равен 0.168 или 0.144, соответсвенно.
Сравнив значения R и Rfree, так как они не сильно отличаются,можно сделать вывод, что модель не
переобучается, они маленькие, что говорит о том, что модель соответствует
данным. Средний B-factor равен 11.3, т.е. небольшой, что хорошо.
Для всей структуры, включающей цепи А и В, маргинальный остаток по карте Рамачандрана всего один (глицин-9 цепи А) из 274 анализированных, т.е. согласно округлению, представленному
в отчете о валидации 0% или 0.4%.
В структуре белка есть остатки с низким критерием качества геометрии и плохим совпадением с ЭП. Для цепей А и В остатков с плохой геометрией
примерно 5-6 %, а с несоответсвием ЭП 6-8%, причем некоторые такие остатки находятся на С-конце, где у данного белка располагался гис-тег.
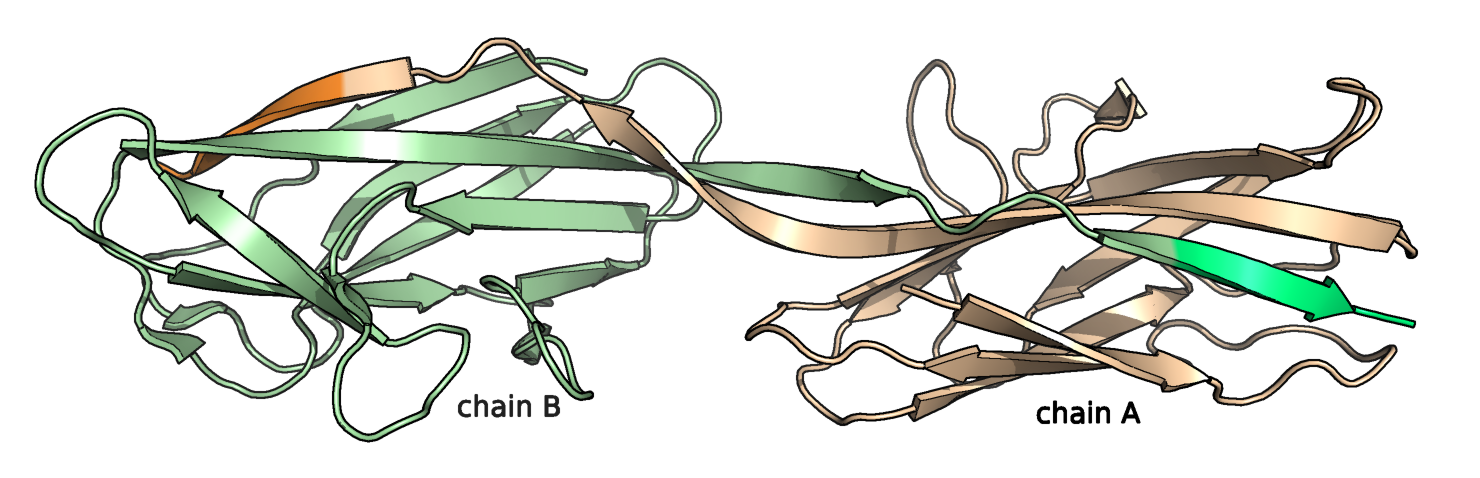
Рис.2. 2AXW, цепи А и В (мономеры димера), С-концевые гис-теги выделены другим оттенком.
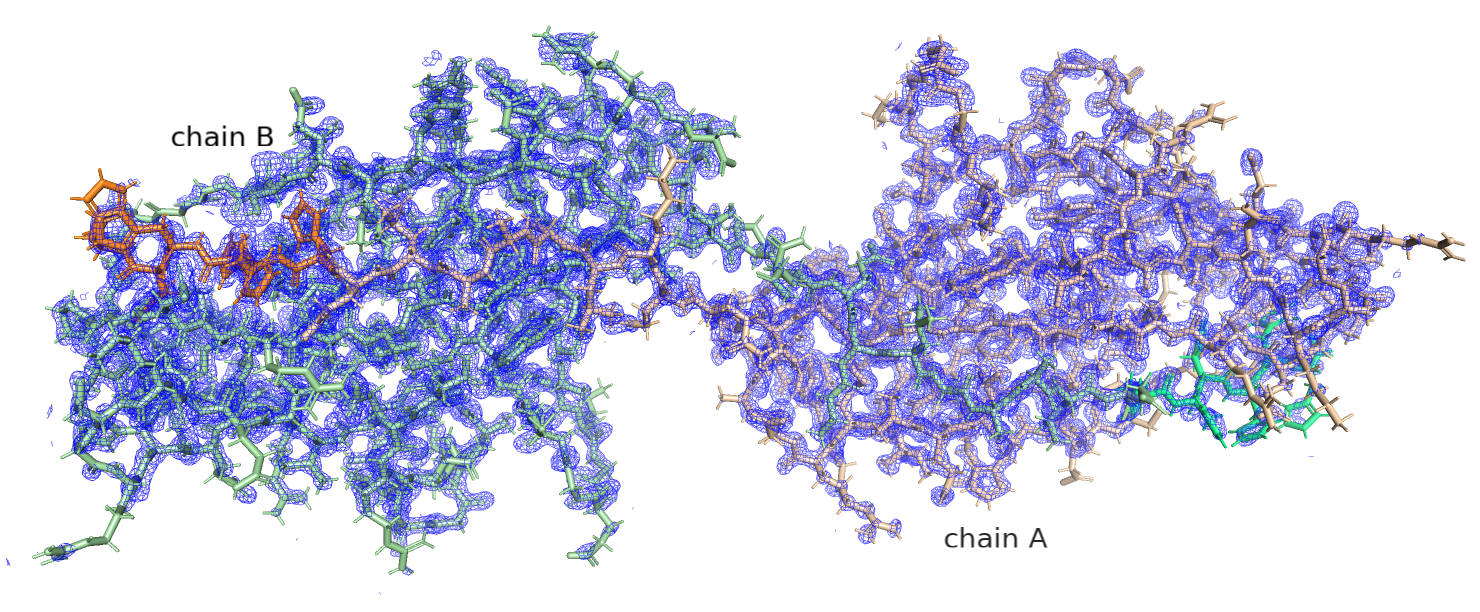
Рис.3. 2AXW, цепи А и В, показана электронная плотность при уровне подрезки 1.5
Рис.3. 2AXW, цепи А и В, показана электронная плотность, увеличенный участок с остатками, плохо вписывающимися в ЭП
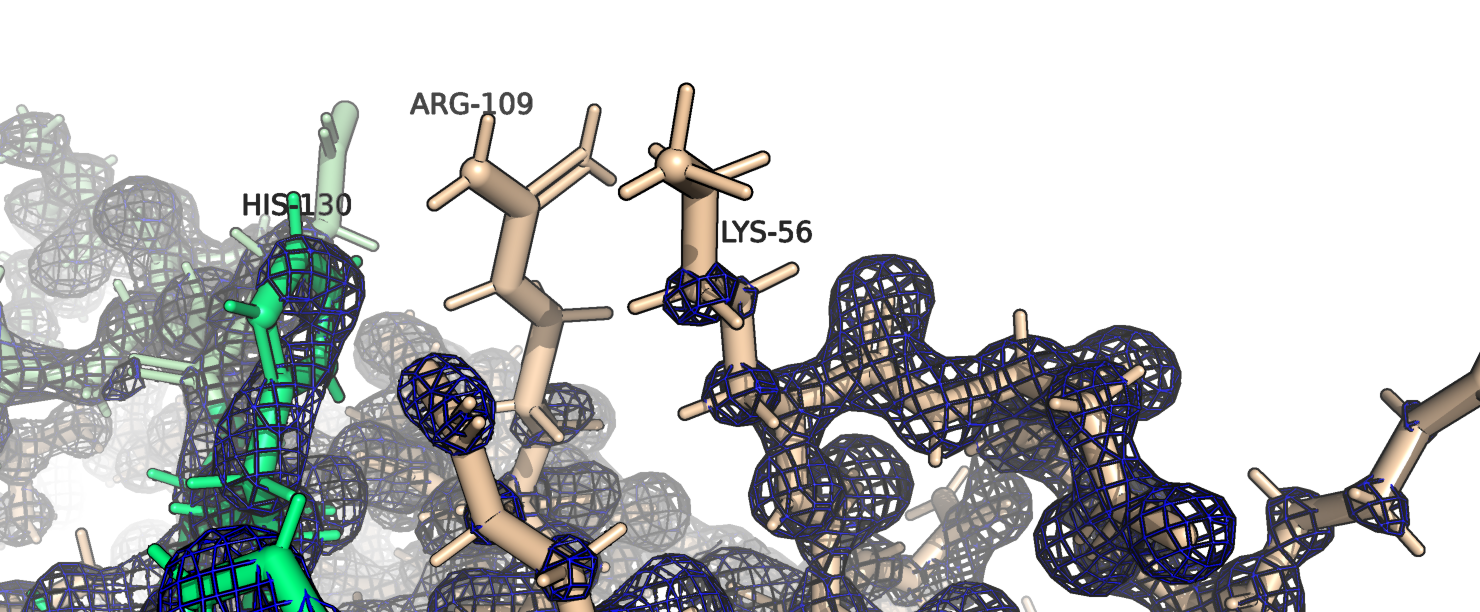
На рис. 3 видно, что лизин и аргинин, которые плохо вписываются в ЭП, находятся рядом с той областью,
в которой происходит димеризация (зеленый гистидин на рисунке - от другого димера).
Можно предположить, что если бы мы изучали взаимодействие не в форме гомодимера, а in vitro функцию, эта область была бы нам очень интересна.
Также такие остатки есть в начале белковой цепи, это, в основном, те остатки, которые, согласно Uniprot, являются сигнальным пептидом,
структура которого важна для распознавания другими ферментами клетки.
Задание 2
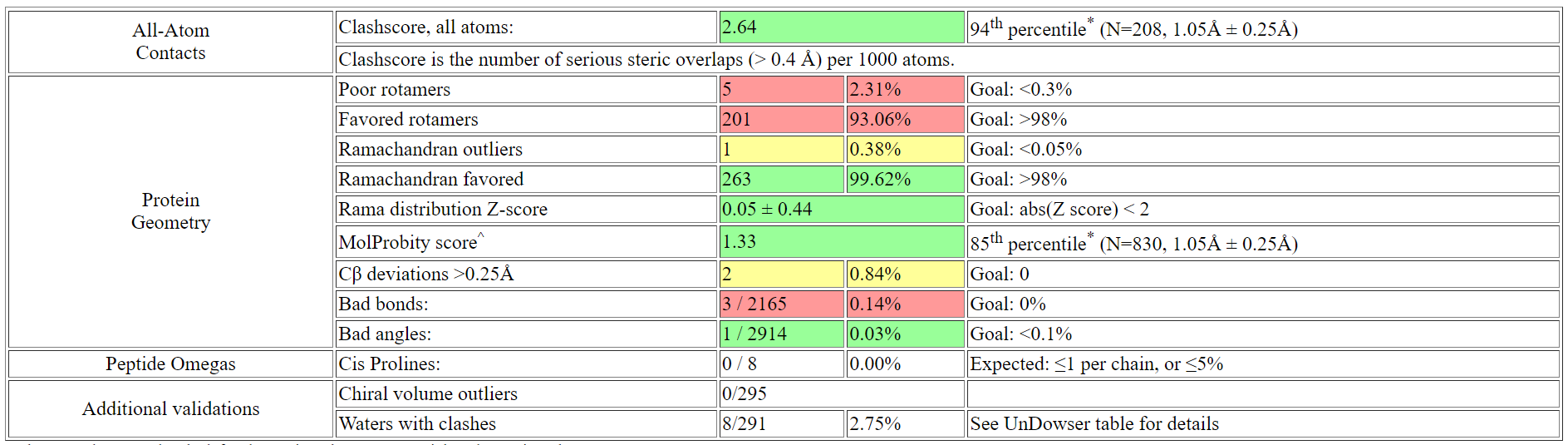
С помощью MolProbity я проанализировала качество структуры без добавления водородов. Полученные результаты можно увидеть
здесь. Просмотрев сводную таблицу в выдаче анализа геометрии MolProbity, можно сднлать вывод, что
в данной структуре многогвато остатков с нехарактерными положениями. Содержание редких ротамеров составило 2.31%, что почти на порядок выше 0.3%.
В структуре также 1 аутлаер карты Рамачандрана, но с учетом длины белка, это тоже выше нормы почти на порядок (0.38%).
Кроме того, обнаружено 3 "плохих связи", которых не должно быть.
Если более детально посмотреть на анализ, можно заметить, что оснавная часть аутлайеров и редких ротамеров находятся
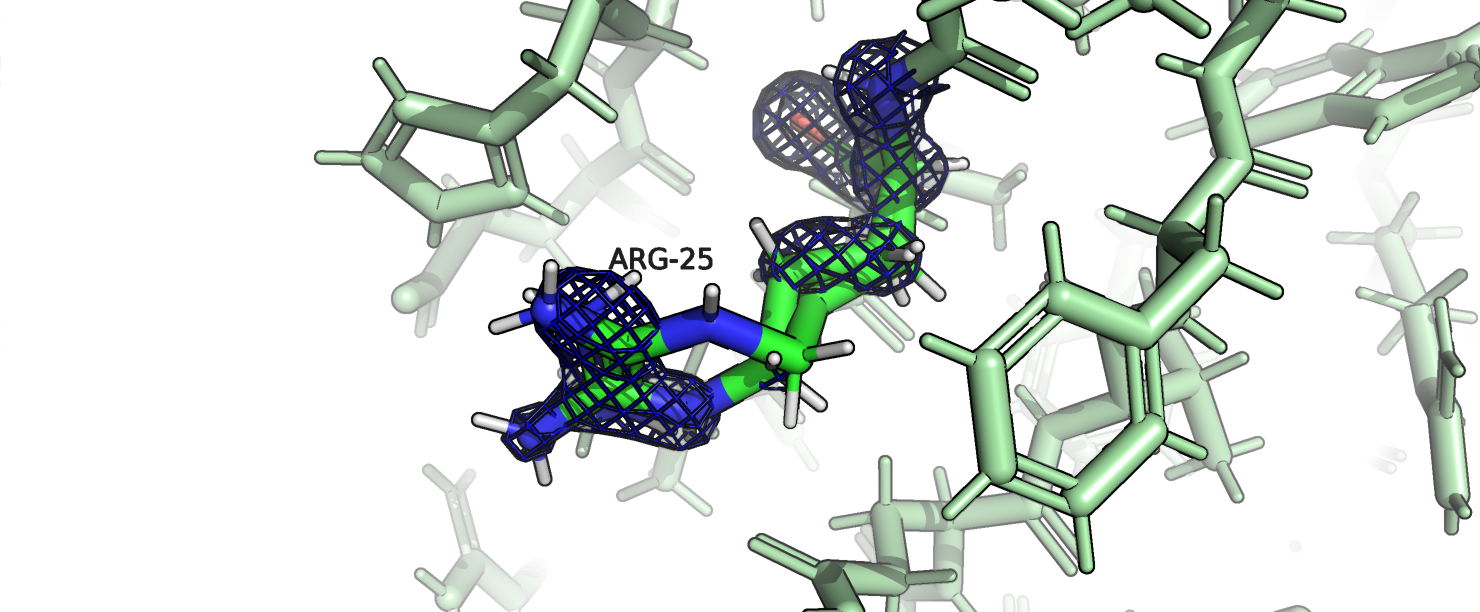
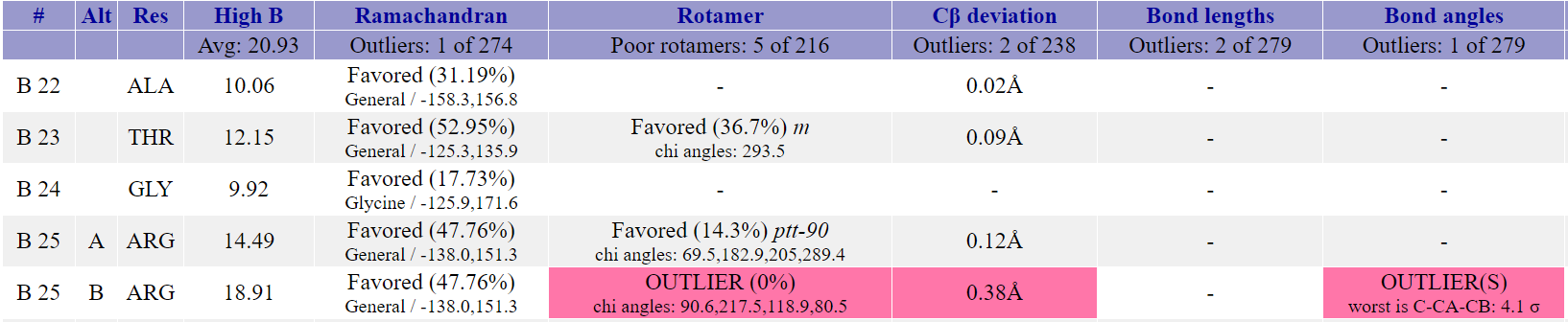
в начале полипептидной цепи. Самый нестандартный остаток - это аргинин-25 цепи B. У этого редкого ротамера также превышен Cβ deviation и угол C-CA-CB.
На рис. 5 изображен этот остаток, можно заметить, что в структуре он имеет два альтернативных конформера, причем нестандартные характеристики относятся к
конформеру B.
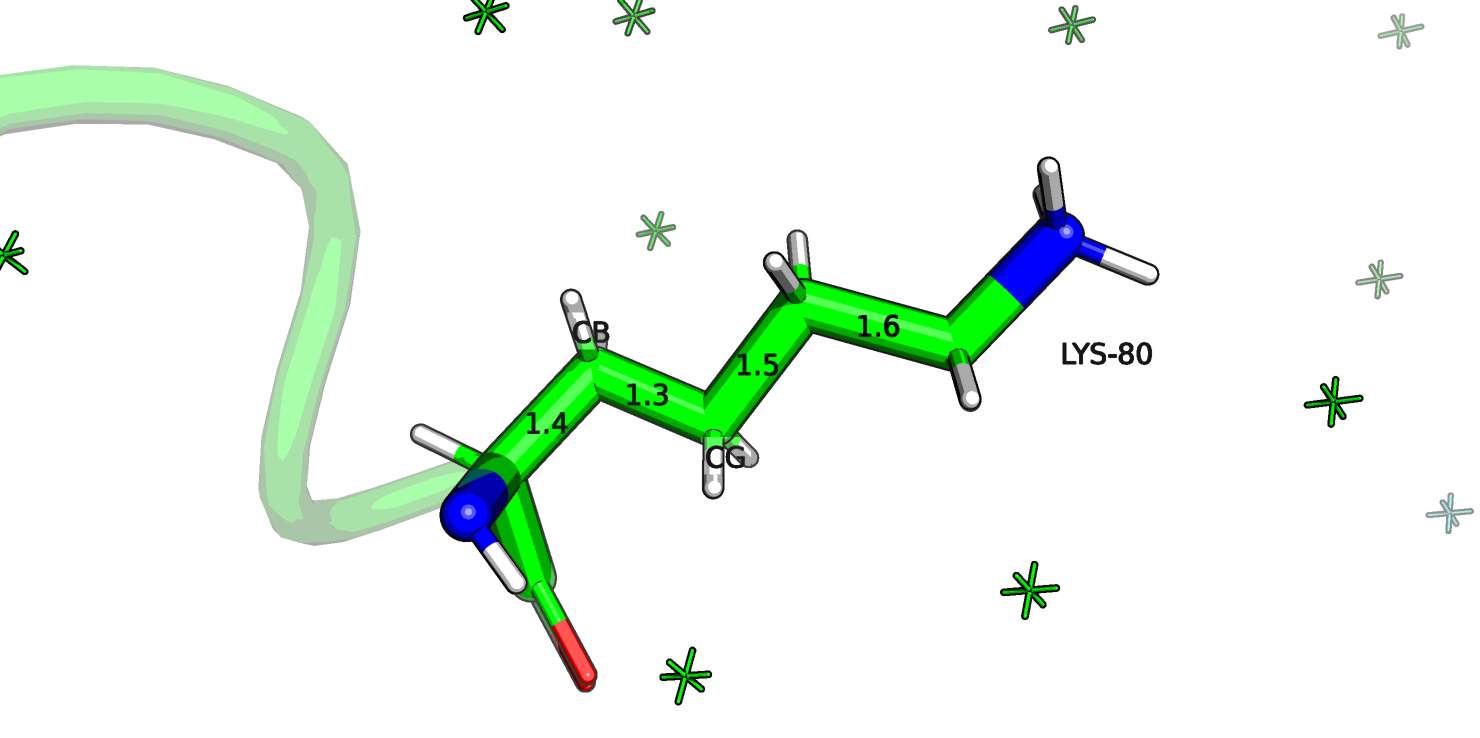
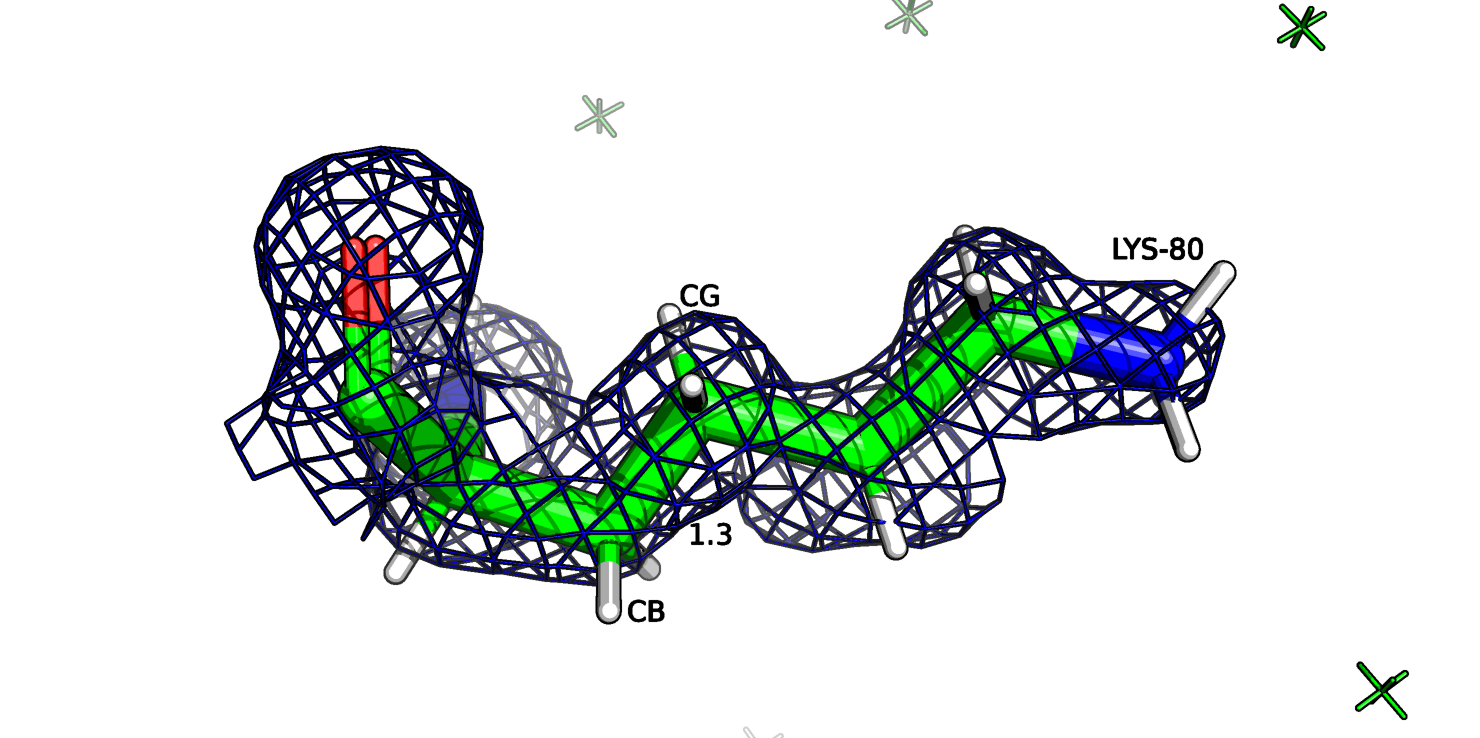
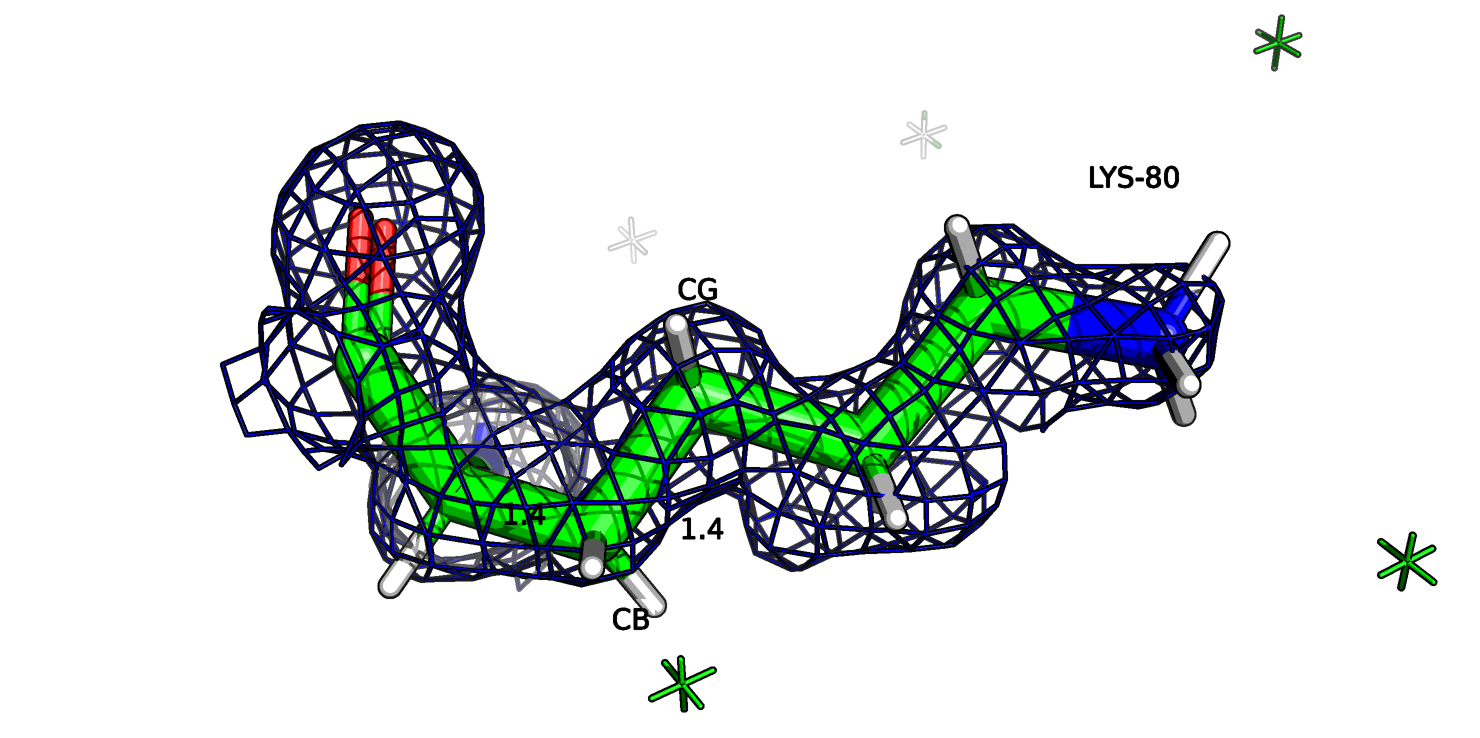
Рассмотрим также остаток лизина-80 цепи А (рис.16а). Этот остаток является аутлаером по длине CB-CG связи (5.7 σ), на рисунке длина связи отмечена 1.3 ангстрем.
Если посмотреть на ЭП этого остатка, то в целом, атомы соответствуют ЭП, но вокруг СВ атома плотность не такая округлая как у других атомов (рис. 16b) и центр атома можно
немного сдвинуть вручную в сторону, и тогда длина связи СА-СВ и СВ-CG будет 1.4 ангстрем (рис. 16с).
Рис.5. 2AXW, Аргинин-25, выделен зеленым
Рис.6. Результаты анализа геометрии структуры
Рис.16а. 2AXW, Лизин-80
Рис.16b. Лизин-80 и ЭП
Рис.16c. Лизин-80 и ЭП со сдвинутым ценром атома СВ
В конформер остатка валина-21 цепи B является аутлаером по значению С-бета отклонению (0.35 А). Возможно это связано с тем,
что ЭП двух конформеров накладывается друг на друга и центры атомов могут быть определены не так точно.
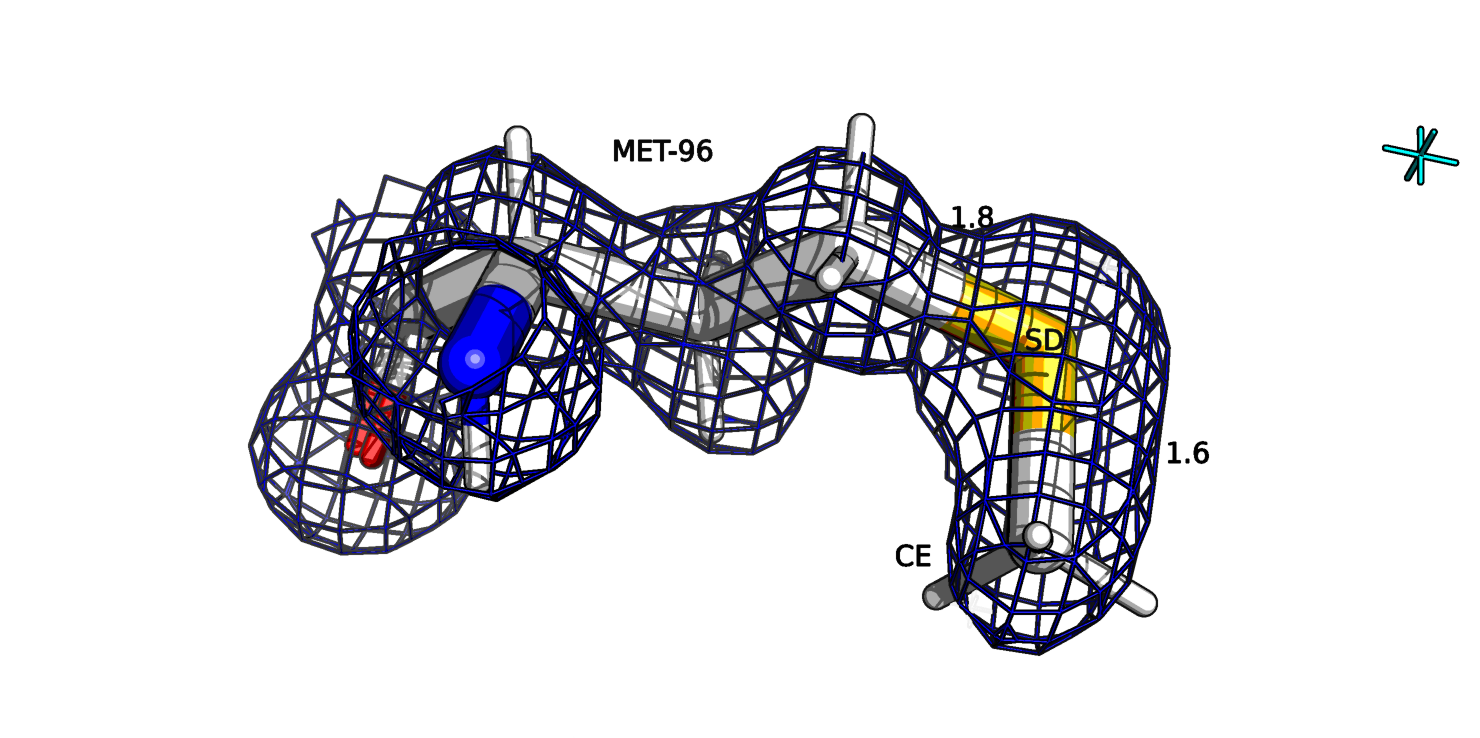
Рассмотрим остаток метионина-96 цепи В. Обычно вроде как длина связи сера-СЕ в метионине равна примерно 1.8 ангстрем.
У данного остатка SD--CE: 6.9 σ, 1.6 А. (рис. 17). Возможно это связано с тем, что СЕ атом находится как бы на конце остатка и ЭП больше смещена с сере и
поэтому центр атома определился ближе к сере.
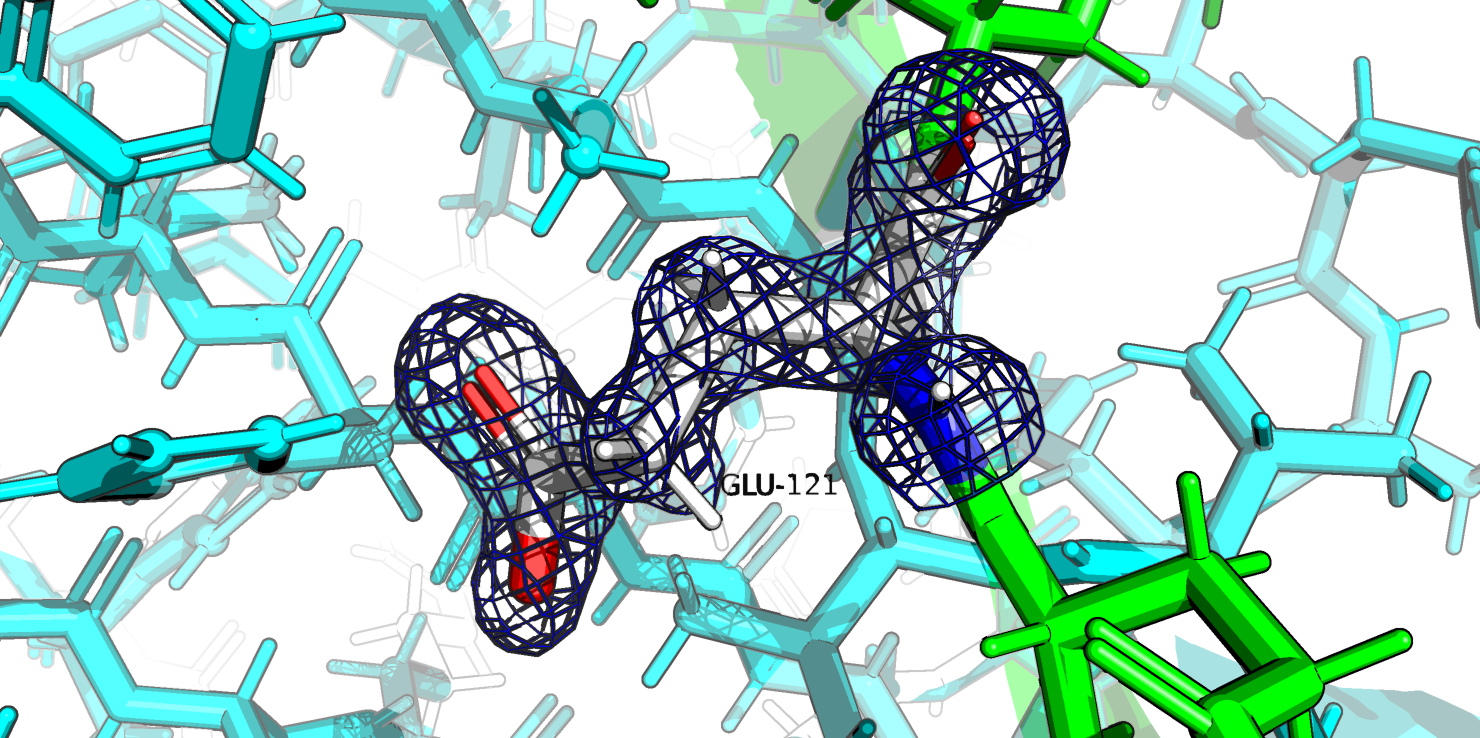
Глутамат-121 цепи А является редким ротамером с углами хи: 312.4,242.4,319.4 . То, что он является ротамеров возможно связано с тем, что он ображен внутрь белка
и такая конформация стабилизируется взаимодействиями с дргуими остатками.
Примечательно, что в структуре нет маргинальных молекул воды.
Рис.16а. 2AXW, метионин-96
Рис.16b. Глутамат-121
\
Затем я провела анализ all-atom contacts and geometry с добавленными водородами. При
добавлении водородов MolProbity также производит инверсии His/Asn/Gln, так что качество
модели могло улучшиться. Однако данные, представленные в таблице суммароной статистики не изменились. В таблице с Multi-criterion chart
появился новый столбец Clash, который показывает, когда недонорно-акцепторные атомы перекрываются более чем на 0,4 Å,
что считается как серьезное столкновение (clash), которое включается в отчетную оценку столкновений. Clashscore по всем атомам равен 2.64 (хороший).
Рис.7. Результаты анализа геометрии структуры по всем атомам с водородами
Задание 3
В целом, в структуре, конечно, есть ошибки, но они в основном затрагивают функционально незначимые остатки. Высокое разрешение позволяет
предполагать небольшое количество ошибок в интересующих нас участках.
Часть тех остатков, которые являются редкими ротамерами или имееют какие-то отклонения,
являются одним из альтернативных конформеров. Например, глицин 43 и аргинин-25, валин-21 цепи В, валин-52 цепи А; в случае аргинина-63 цепи А оба альтернативных
конформера имееют большой clash, но с разными атомами.
У белка нет функционально значимых кофакторов, так что точность расположения
иона хлора и глицерола в структуре не принципиально важна. В полученной структуре на С-конце находится гексагистидиновый тег, который не
помешал образованию димера, можно предположить что при взаимодействии с другими белками в клетке, конформация белка не отличается от полученной методом РСА.
Атомы в большинстве случаев хорошо описываются электронной плотностью, кроме тех остатков, которые сильно выступают на поверхности глобулы.
Так как данный белок не фермент, у него нет остатков активного центра, расположение которых нам было бы очень важно узнать.
Можно сказать, что качество структуры достаточно хорошее.
Задание 4
В этом задании я сравнивала описанную выше структуру PDB с переделанной в моделью в PDB Redo. На следующем рисунке представлена таблица с метриками
переделанной модели.
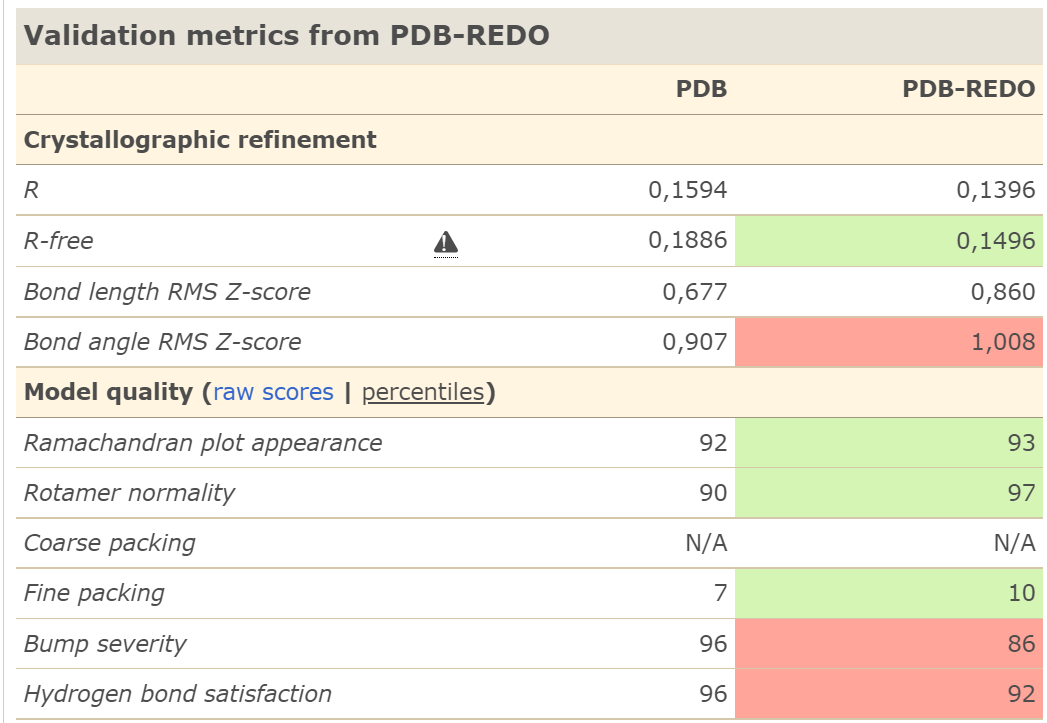
Рис.8. Валидация переделанной модели 2AXW в PDB Redo
Сравнивая результаты валидации, можно заметить, что значения R-фактора и R-free,
представленных как значения из
PDB немного отличаются от тех, которые были представлены в отчете о валидации ранее. Причем,
R-free was considered biased, the estimated unbiased R-free was used. В переделанной модели эти параметры были уменьшены.
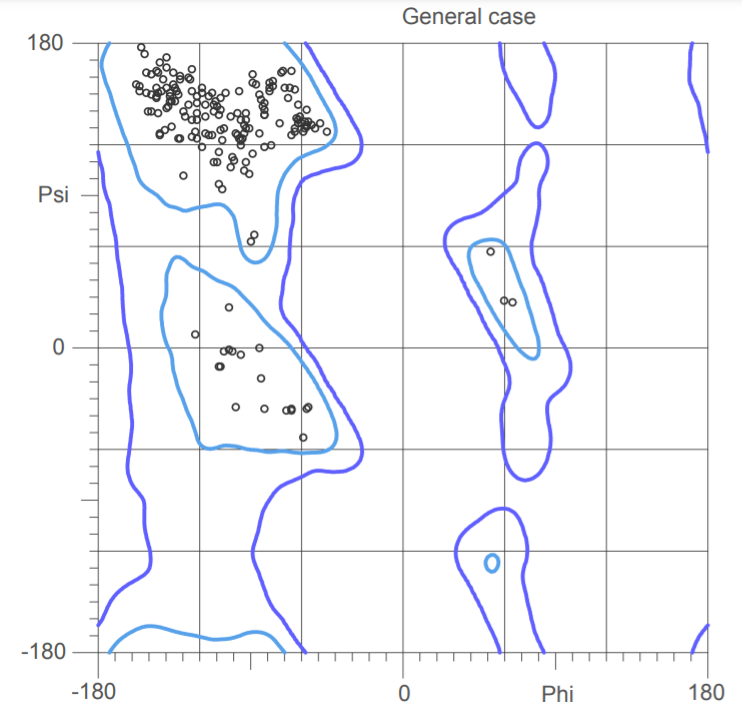
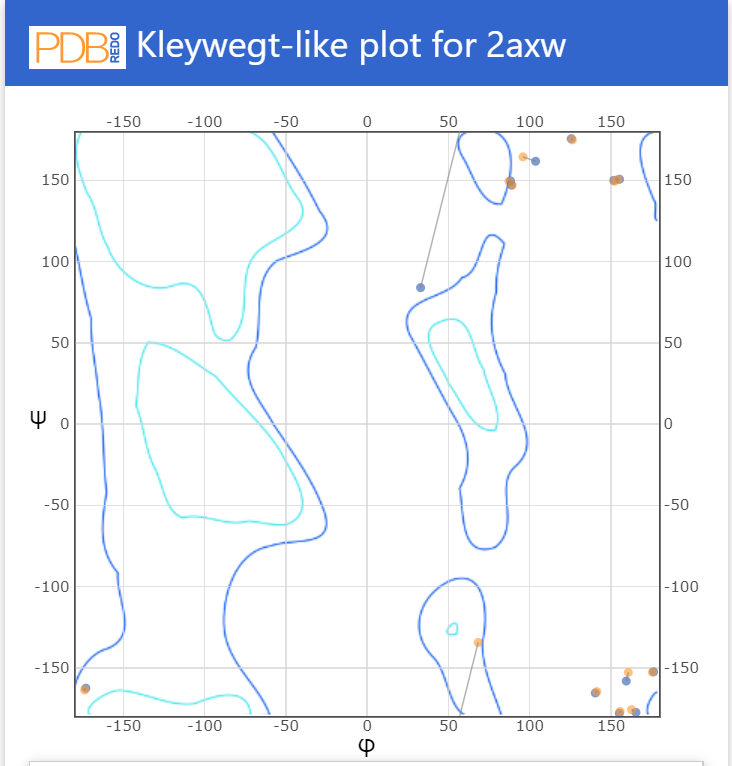
Качество карты Рамачандрана увеличилось на 1, сравнение карт можно увидеть на рис. 9 и 10.Нормальность ротамеров с 90 до 97. Показатель "прекрасной укладки" вырос с 7 до 10,
и "серьезность неровностей" упала с 96 до 86, хотя осталась плохой.
Рис.9. Карта Рамачандрана из MolProbidity
Рис.10. Карта Рамачандрана PDB Redo
Существенные изменения модели. Было изменено 5 ротамеров и удалено 15 молекул воды, одна инверсированная пептидная связь.
В итоге 3 остатка стали хуже соответствовать ЭП, но 153 остатка стали лучше.
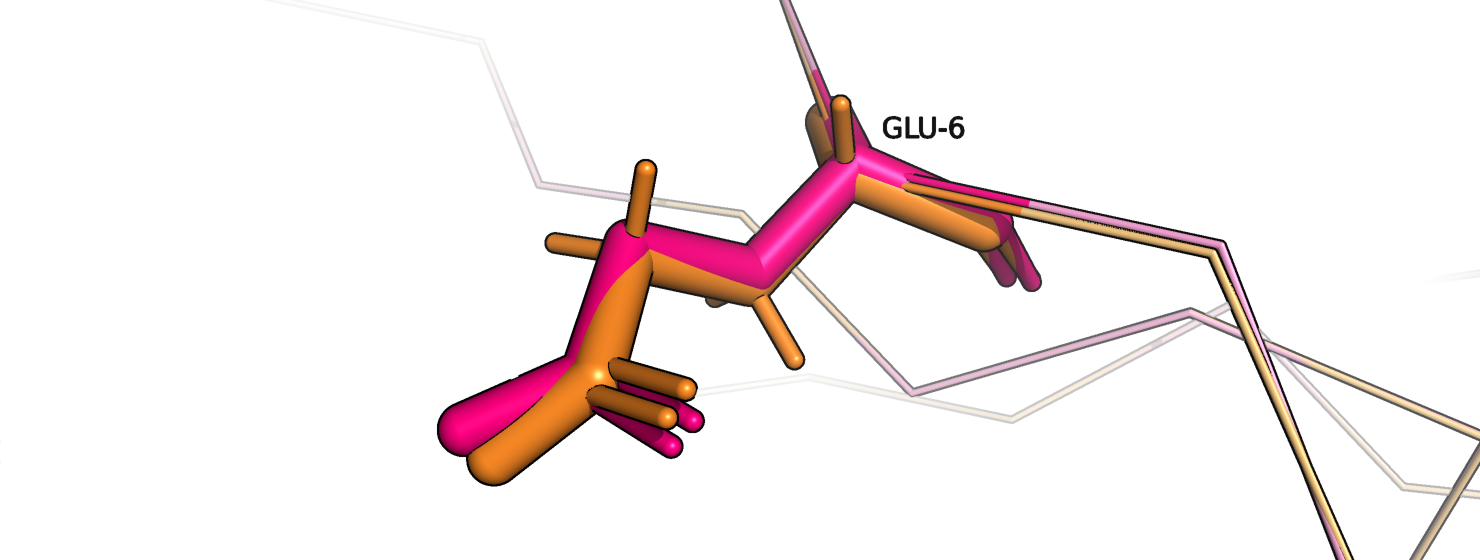
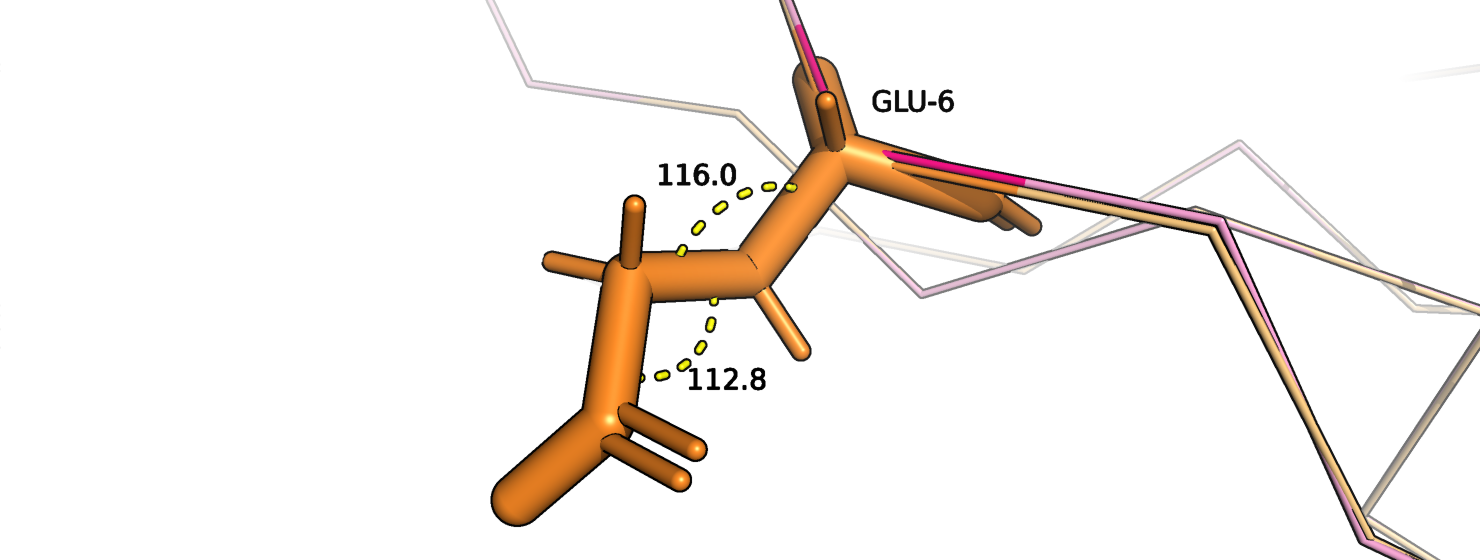
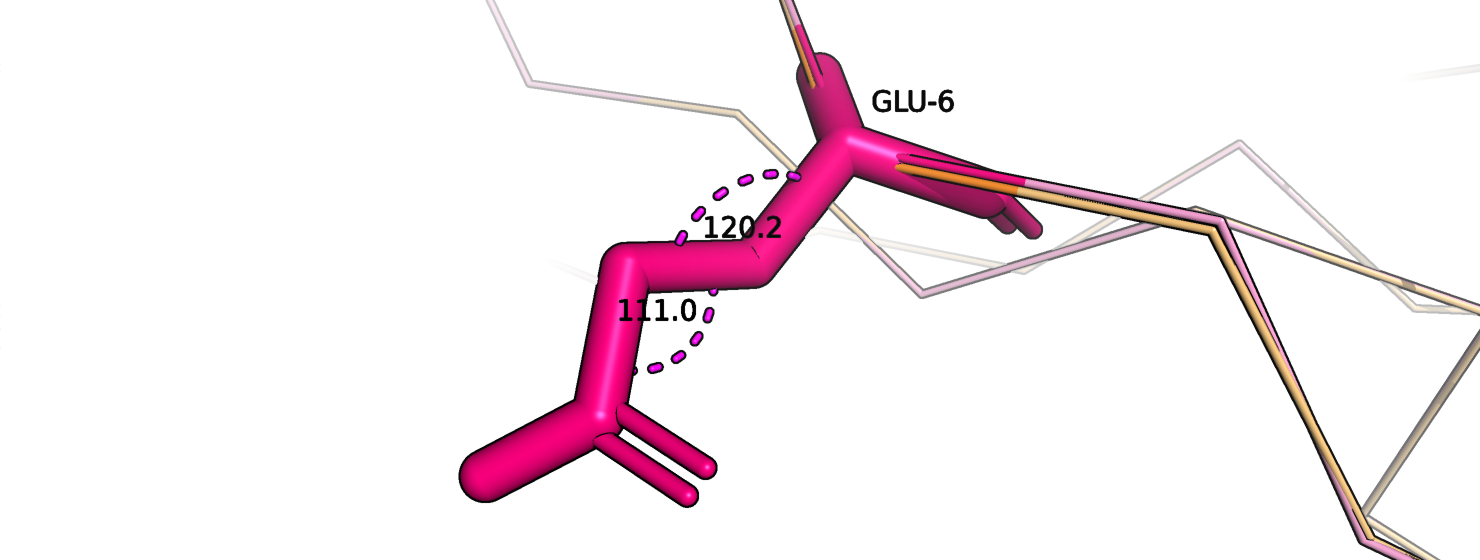
Рассмотрим на примерах. На рис. 11 изображен остаток глутамата-6 цепи А, который относится к сигнальному пептиду. Оранжевым покрашена
исходная струтктура, розовым - новая модель. Хи-углы стали получше (рис. 12-13).
Рис.11. Глутамат-6, цепь А. Оранжевый - исходный, розовый переделанный
Рис.12. Глутамат-6, цепь А. Исходная структура
Рис.13. Глутамат-6, цепь А. Переделанная модель
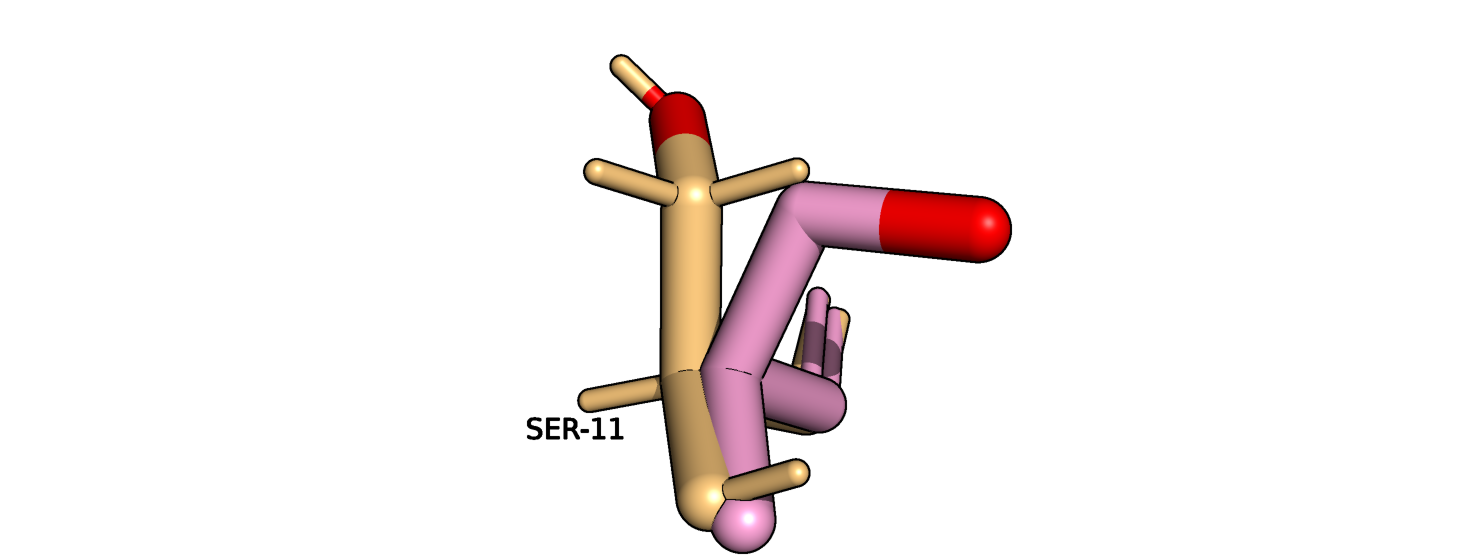
Наиболее сильному улучшению подвергся остаток серина-11 цепи B (рис. 14). Однако его положение
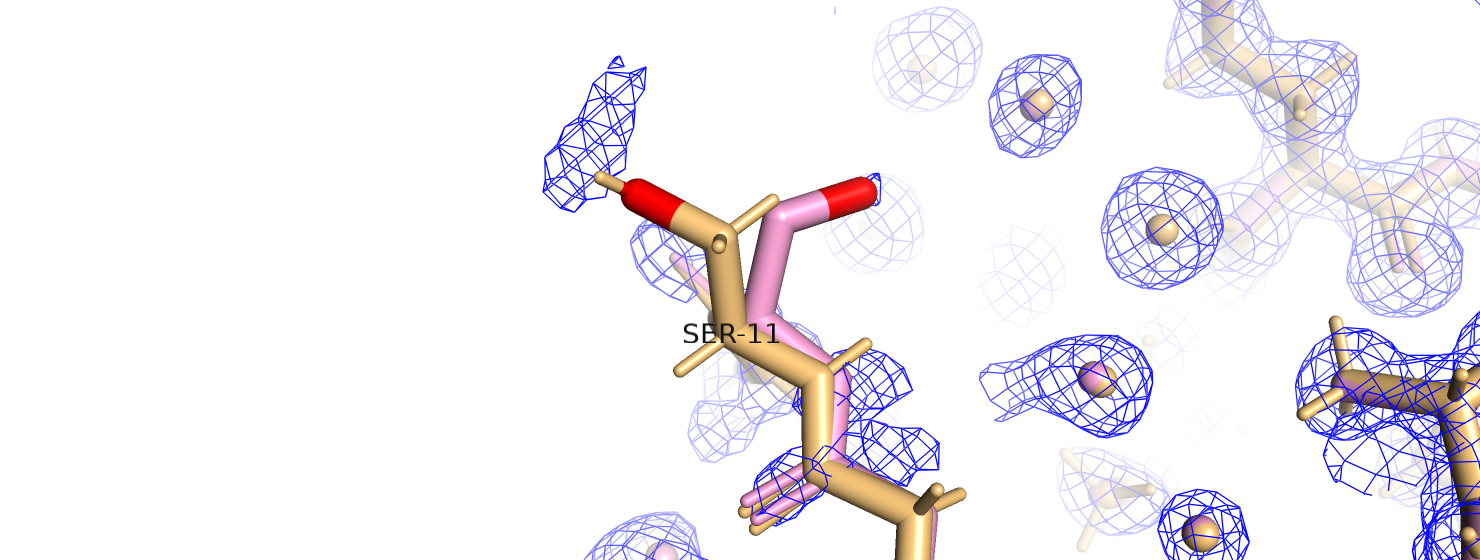
не очень то хорошо вписывается в ЭП (рис. 15). Если доверять такому улучшению, так как этот остаток тоже находится
в составе сигнального пептида, оно может позволить изучить расположение остатков, распознающихся другими белками. Хотя, в принципе,
в исходной структуре данный остаток не был маргинальным. С другой стороны, маргинальный остаток аргинина, описанный выше, при переделывании не улучшился, а немного ухудшился.
Таких остатков, которые бы существенно ухудшились, в новой модели нет.
Рис.14. Серин-11
Рис.15. Серин-11 вписанный в ЭП
Таким образом, если говорить про всю переделанную модель, она, конечно, улучшилась, но незначительно. Каких-то
радикально важных изменений внесено не было, структура была хорошая, после переработки стала еще получше. Наибольшим улучшениям подверглись молекулы воды.
Если проводить изучение данного белка, принципиальной разницы между моделями нет, поэтому можно пользоваться как переделанной, так и исходной.