Изучение особенностей археи Halanaeroarchaeum sulfurireducens на основе данных генома
Соколов Сергей Ильич
Факультет биоинженерии и биоинформатики, Московский государственный университет им. М.В. Ломоносова, Ленинские горы, д. 1, стр. 73, 119234, Москва, Россия
Резюме
Предпосылки: исследования гиперсолёных озер в Кулундской степи (Алтай, Россия) 2009-2013 годов привели к открытию нового вида галоархей с ранее несвойственным царству архей метаболизмом [1]. В данном мини-обзоре я буду проводить анализ некоторых данных о геноме этого организма.
Введение
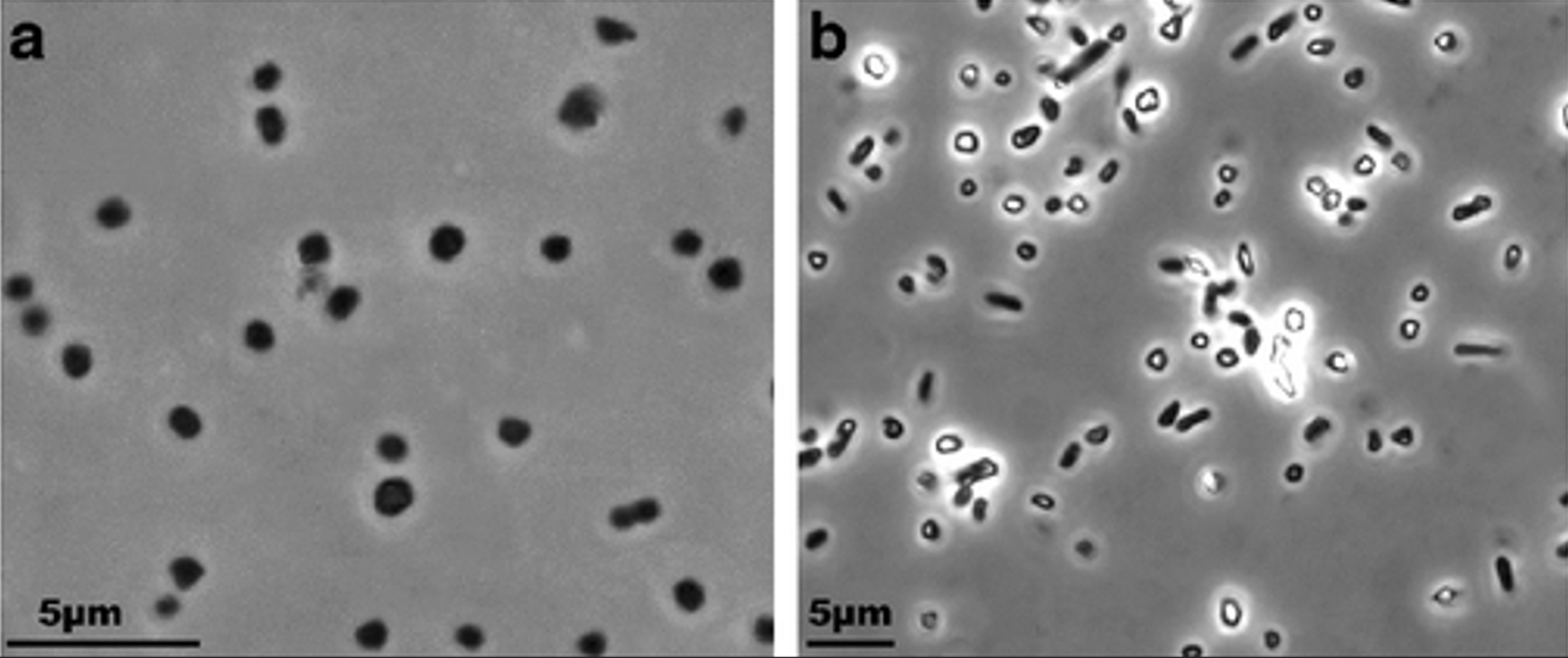
Вид является единственным описанным в своём роде, представляет из себя анаэробный организм, неподвижный, плоский коккоид или досковидную палочку (рис. 1). Метаболизм чрезвычайно узкий: в качестве донора электронов/источника углерода используются ацетат и пируват, а в качестве акцептора-элементарная сера; в качестве источника азота используется аммоний. Архея была обнаружена в рассолах и аэробных сульфидных поверхностных отложениях (2-10 см), полученных из гиперсоленых хлоридно-сульфатных озёр. Оптимальный рост происходил в присутствии 4М (Моль/Л) NaCl и в диапазоне от 3 до 5 М при температуре 37-40 °С. Удивительно то, что в одной из культур (штамм HSR2) наблюдались такие восстановленные соединения серы, как 𝐶𝑆2, 𝐶𝐻3𝑆𝐻, которые никогда не встречались у известных серовосстанавливающих прокариот [1]. Таксономическое положение археи: Домен: Archaea, Царство: Euryarchaeota, Класс: Halobacteria, Отдел: Halobacteriales, Семейство: Halobacteriaceae, Род: Halanaeroarchaeum, Вид: Halanaeroarchaeum sulfurireducens [1]
Рис. 1. Изображение Halanaeroarchaeum sulfurireducens, выращенных в ацетате (а) и пирувате (b) в качестве доноров электронов и элементарной сере в качестве акцептора электронов [1]
Методы
При изучении археи использовались нижеперечисленные платформы и их средства:
2. Интерпретатор команд Bash, в котором при помощи команды infoseq *_cds_from_genomic.fna -nocolumns -out cds.csv, я создал файл с кодирующими последовательностями, а затем через подключение к серверу по протоколу sftp скачал его и таблицу генетических особенностей, полученную из банка генов: GCF_001305655.1_ASM130565v1_feature_table.txt. Все вышеперечисленное было импортировано в Google таблицы.
3. Google таблицы (Excel) (см. сопроводительные материалы, п.1), где и проводился анализ генома с использованием формул и функций (последние я привел ниже для каждого задания):
3.1: MAX, MIN, COUNTIFS
3.2: COUNTA
3.3: COUNTIF
3.4: MAX, MIN, COUNTIF, COUNTIFS
3.5: COUNTIFS
3.6: -
3.7: MAX, MIN, COUNTIFS
Результаты
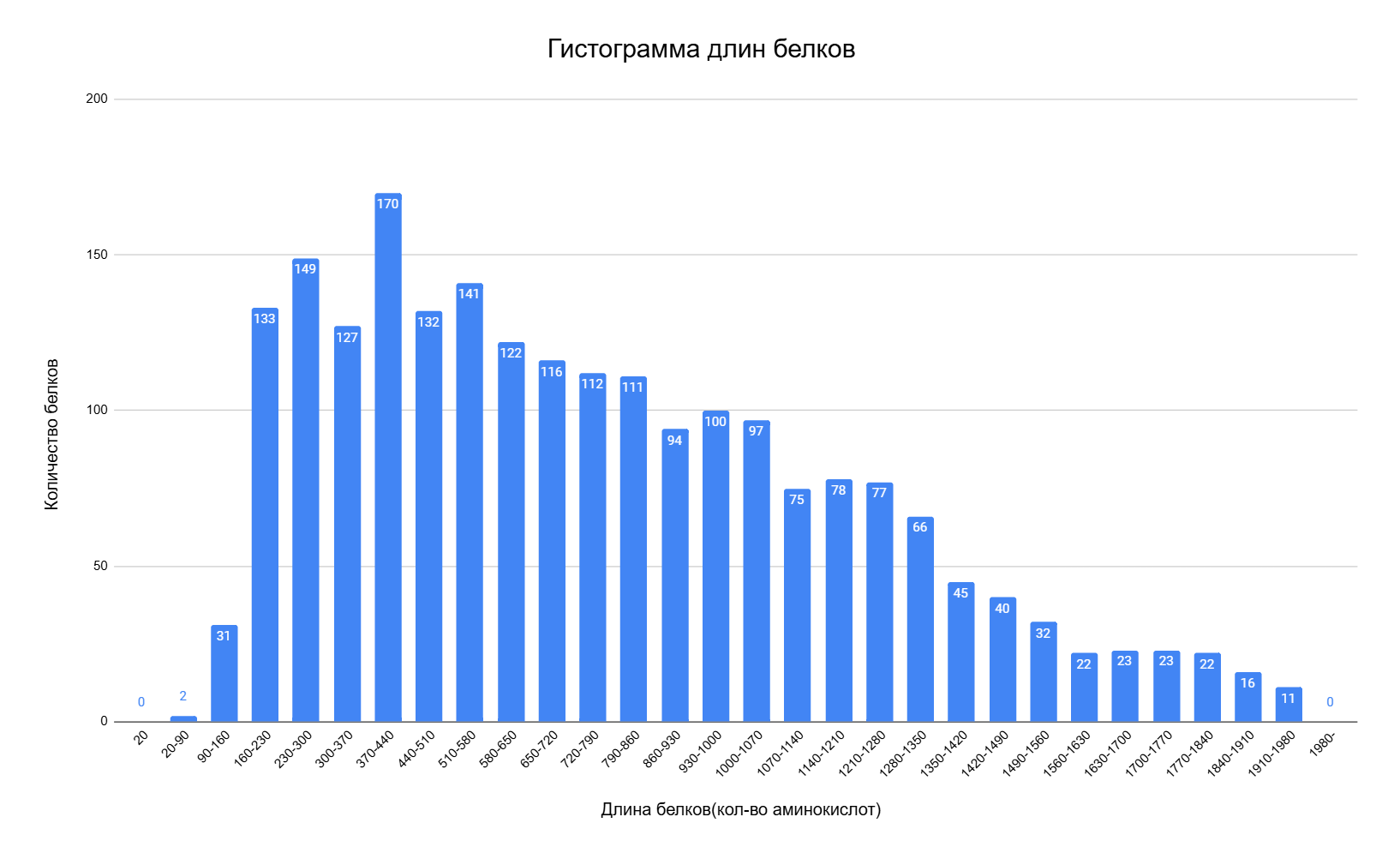
Гистограмма длин белков, закодированных в геноме археи Halanaeroarchaeum sulfurireducens
Из гистограммы можно сделать некоторые выводы о белках археи (рис. 2):
1. Степень полимеризации молекулы. Чем большим количеством аминокислот закодирован белок, тем, вероятнее, сложнее его пространственная структура. Пространственная же структура определяет функции белка, количественно или качественно [2].
2. На гистограмме отсутствуют пробелы, однако наблюдаются пики, т.е. определённые белки с длинами от 160 до 860 являются наиболее функционально значимыми для археи. Белки с большей длиной встречаются реже, и это естественно: их сложнее синтезировать.
3. Наиболее распространены белки длиной 370-410 аминокислот – 170 единиц. Белки с длиной менее 20 и более 1980 аминокислот не встречаются.
В среднем белки эукариотических организмов длиннее бактериальных, а последние, в свою очередь, длиннее архейских. Этот факт можно интерпретировать как эволюционную тенденцию к увеличению размера белка, которая свидетельствует о слиянии однофункциональных белков в расширенные многофункциональные [2].
Рис. 2. Гистограмма длин белков
Длина белка часто связана с его функцией. Короткие (пептиды) в основном выполняют простые, но специфические функции, например, сигнальные или регуляторные белки. Длинные имеют в своём составе
несколько доменов (домен – компактная, независимо сворачивающаяся часть белка, связанная определённой функцией), и выполняют множество функций, они более
склонны к различным мутациям, т.к. гены, кодирующие такие белки, по своей величине им не уступают, следовательно, вероятность случайной мутации увеличивается [3]. Нет определённого числового значения, при котором белки начинают менять свою
пространственную структуру, и она зависит далеко не от количества аминокислот в их составе, но приблизительно эти значения таковы: от 2-40 аминокислот – короткий пептид; от 40 до 100 – небольшие белки; от 100 до 1000 – длинные белки; белки длиной более 1000 аминокислот считаются очень длинными.
Сравнение участков генов (CDS)
CDS – Coding Sequence. Это участки генов ДНК или РНК, которые кодируют последовательность аминокислот белка. Собственно, рибосома считывает именно этот участок в процессе трансляции. Соотношение неактивных CDS от общего количества CDS (CDS without protein от CDS) может указать на ряд фактов о геноме организма [4-5]:
1. Если количество CDSwp довольно высоко, (от 20%) это означает, что геном не оптимизирован, и содержит много нефункциональных участков. Это могут быть псевдогены, регуляторные элементы, а также следствия мутаций. В процессе эволюции гены могут становится нефункциональными, но не удалёнными из организма; горизонтальный перенос между несколькими прокариотами также ведёт к
увеличению данного соотношения, поскольку не все гены одного организма подходят к другому. Имеет место процесс псевдогенизации.
2. Если количество CDSwp низкое, то большинство участков приводят к синтезу рабочих белков. Организму не приходилось мутировать множество раз, переносить гены от другого организма, а также менять род жизнедеятельности.
Количество таких участков было подсчитано в моих электронных таблицах. CDSwp – 30, общее количество CDS – 2282. Следовательно, доля CDSwp составляет 0,013, т.е. 1,3%.
Сравнение репликонов в архее и их состава
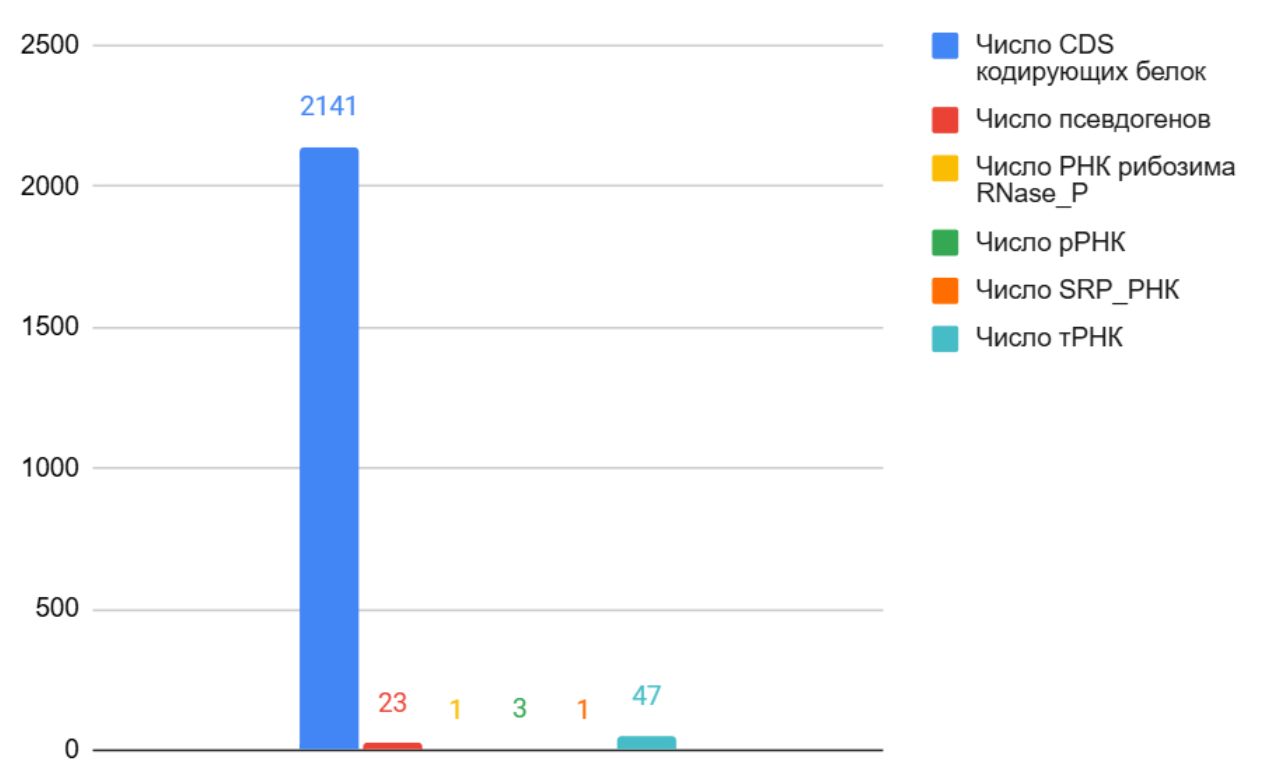
Репликонами принято называть кольцевые молекулы ДНК. В составе таблиц они находятся в столбце Chromosome, самый большой репликон обозначается пустым промежутком. Сравнивая их состав, т.е. какие гены с какими функциями они содержат, можно сказать, собственно, о функциях самого репликона. Рассмотрим репликоны данной археи, их всего два, в порядке уменьшения (рис. 3-4).
Рис 3. Гистограмма количества генов и их функций на наибольшем репликоне
Наибольший репликон (рис. 3) содержит гены со всеми функциональными единицами. Т.к. роль белков в организмах невероятно обширна, генов, их кодирующих, преобладающее большинство. Гены, несущие псевдогены, более не функциональны и не представляют особого интереса, их доля составляет 0,01, т.е. 1%. Отдельно стоит обсудить гены, несущие различные виды РНК, а именно сами РНК:
1. RNase_P_RNA: данная РНК является каталитической частью рибонуклеопротеина RNase P (рибонуклеаза P). RNase P – это рибозим, т.е. фермент на основе РНК (один из немногих), который участвует в процессе доработки РНК, а именно в созревании тРНК. Рибозим чрезвычайно консервативен во всех организмах его содержащих, т.е. выполняет строго определённую функцию. Сама РНК является одной из древнейших, т.к. без неё невозможно синтезировать тРНК, которая необходима для трансляции белков [6].
2. rRNA: такие РНК входят в состав рибосом, составляя их каркас, участвуя в формировании пептидных связей (Большой субюнит) и расположении мРНК и тРНК для успешной трансляции (Малый субюнит). Как и предыдущая РНК, чрезвычайно консервативна.
3. SRP_RNA: РНК представляет собой компонент сигнального распознающего комплекса (Signal Recognition Particle), который участвует в процессе транспортировки белков через клеточную мембрану. Сигнальные пептид обычно располагается на конце белка, и указывает, что белок должен быть транспортирован через какую-либо мембрану. Данная РНК способствует связыванию комплекса с рибосомой, синтезирующий белок с таким пептидом [7].
4. tRNA: РНК транспортирует аминокислоты к рибосоме, где происходит синтез полипептидной цепи. Каждая тРНК имеет уникальную последовательность для связывания с определённой аминокислотой. На одной из сторон молекулы находится антикодон, комплементарный кодону на мРНК, для точности связывания.
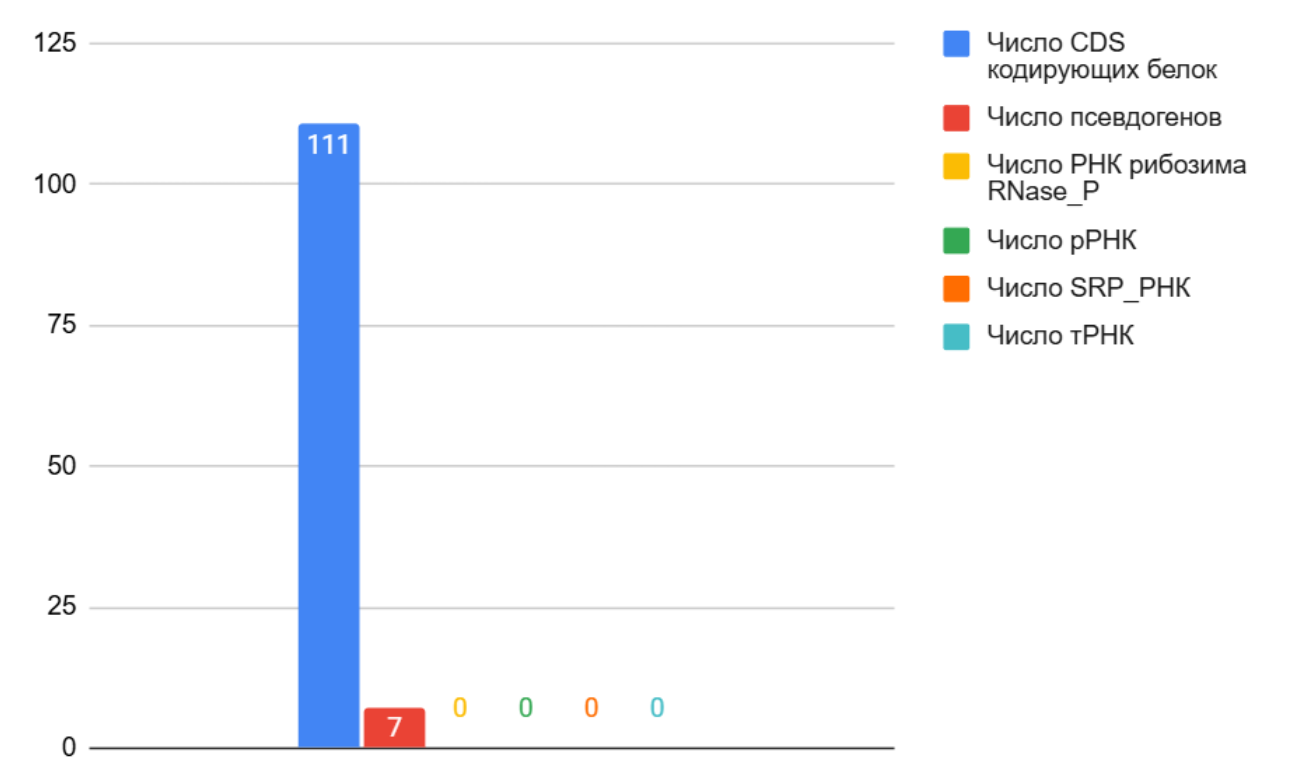
Рис 4. Гистограмма количества генов и их функций на pM27-SA2-01
Следующий и последний репликон (рис. 4) уже имеет название. Из гистограммы видно, что гены, содержащиеся в нём, несут информацию о белках, а также имеются псевдогены, их доля составляет 0,06, т.е. 6%. Такой репликон мог появиться в связи с требованием повышения мобильности и адаптивности археи к различным изменениям окружающей среды, т.е. более эффективно выполнять необходимые задачи без помощи большого репликона, в котором содержится очень большое количество информации [8].
Если сравнить репликоны по составу, очевидно, что репликон pM27-SA2-01 зависит от большого. Во-первых, вероятнее всего, большой репликон кодирует жизненно-необходимые белки, в то время как меньший отвечает лишь за какие-то несколько функций. При этом, все РНК закодированы в большом, следовательно, меньший зависит как минимум по процессам трансляции.
Гистограмма расстояний между участками CDS
Из гистограммы можно сделать несколько выводов (рис. 5):
1. В основном расстояния не превышают 500 нуклеотидов, но есть также участки, расстояние между которыми отрицательное, т.е. считывание начинается с конца другого участка.
2. Расстояний более 8000 нуклеотидов почти не наблюдается, но отдельные большие участки могут представлять определённый интерес, т.к. возможно в процессе эволюции произошло расширение генетического материала, не затронувшее CDS, но изменившее структуру генома [8]
Рис 5. Гистограмма межгенных промежутков
Анализ предоставляет полезную информацию о структуре генома, функциональной организации и структуре организма. Обычно у прокариотических организмов эти расстояния гораздо меньше, нежели у эукариотических, содержащих интроны, регуляторные элементы и некодирующие области в большем количестве.
Близкое расположение участков или их перекрывание может свидетельствовать о наличии оперонов – участков, включающих в себя несколько CDS, но выполняющих очень близкие или зависимые друг от друга функции.
Гистограмма расстояний между пересекающимися CDS
Из гистограммы (рис. 6) видно, что значительная доля перекрываний приходится на значение 4 (то есть перекрываются по 4 нуклеотидам). Это значение может быть обусловлено перекрыванием стоп-кодонов предшествующего участка с старт-кодоном следующего участка. Например, стоп-кодон TGA перекрывается с старт-кодоном ATG. Самое большое перекрывание достигает промежутка от 85 до 90 нуклеотидов.
Рис 6. Гистограмма пересекающихся CDS
Как обсуждалось ранее, перекрывание нуклеотидов между разными участками CDS может свидетельствовать о наличии оперонов. Гены в составе оперона транскрибируются в одну молекулу мРНК. Наличие таких участков помогают клетке скоординированно активировать или дезактивировать определённые биохимические процессы, реализующиеся несколькими белками. Так как прокариотам зачастую необходимо быстро адаптироваться к изменениям окружающей среды, данные участки жизненно необходимы для взаимодействия с окружающей средой с минимальными затратами. Чем больше длина перекрывания между участками – тем более они зависимы друг от друга [9].
Опероны в CDS
Говоря об оперонах, то найти хотя бы несколько таких участков самостоятельно довольно несложно. Необходимо учитывать несколько факторов: расстояние между участками (это может быть и перекрыванием, и просто расположение участков на расстоянии нескольких нуклеотидов друг от друга) и природа того, что этот участок кодирует. Это могут быть белки или РНК, выполняющие функции, относящиеся к одному и тому же процессу в организме.
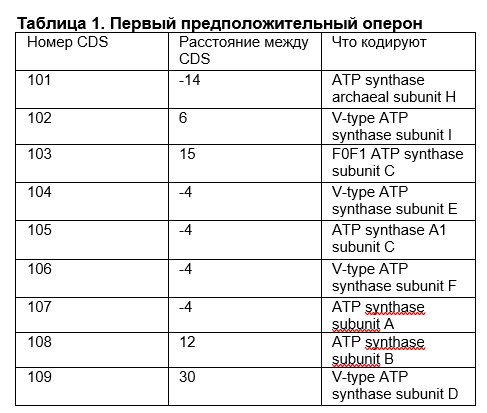
В последующих таблицах (таб. 1-3) представлены предположительные опероны:
1. Гены находятся близко к друг другу, т.е. либо перекрываются, либо располагаются поблизости
2. Все перечисленные гены кодируют субъединицы АТФ-синтазы, которая является мультисубъединичным комплексом (множество субъединиц, объединяющиеся для выполнения определённой биологической функции). Скорее всего, они все экспрессируются одновременно для построения данного комплекса.
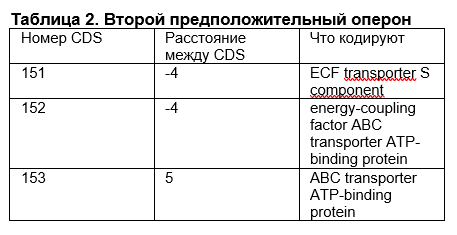
1. Гены снова находятся близко друг к другу
2. Все указанные гены связаны с транспортными белками (ABC – транспортеры и ECF – компоненты), которые выполняют одну задачу – транспорт веществ через мембрану. Вероятнее всего, экспрессируются они все вместе
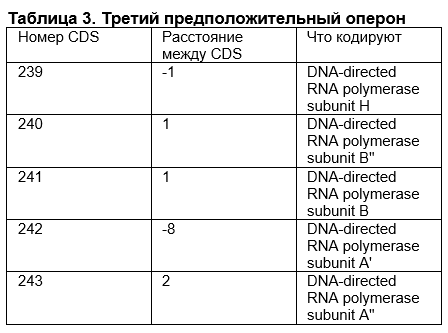
1. Гены вновь расположены близко друг к другу
2. Данные последовательности кодируют субъединицы ДНК-зависимой РНК-полимеразы. Этот фермент участвует в транскрипции – синтезе РНК на матрице ДНК. С большой долей вероятностью, эти участки экспрессируются вместе.
Гистограмма %GC по CDS
Из гистограммы (рис. 7) видно, что содержание GC в CDS велико: в среднем это 60-70%, отдельные участки содержат от 40 до 60%, или от 70 до 80%.
Рис 7. Количество кодирующих последовательностей, распределённых по процентному содержанию GC
Вычисление отношения гуанина и цитозина к общему количеству азотистых оснований необходимо для понимания нескольких важных аспектов об геноме в целом:
1. Высокое процентное содержание GC указывает на большую стабильность ДНК, т.к. связи между гуанином и цитозином более прочные (три водородные связи), чем между аденином и тимином (две водородные связи). Такая молекула более устойчива к разрушению и деформации, разрывам [10].
2. Если процентное содержание GC высокое, это также может говорить об ареале обитания археи, например, о высокой температуре, а как мы знаем, для этой археи условия благоприятны при 37-40 градусах [10].
3. CDS с высоким процентным содержанием GC, вероятнее всего, синтезируют белки, т.к. эти азотистых основания влияют на экспрессию генов, т.е. на их активность при передаче генетического материала в виде аминокислотных последовательностей, а также уменьшают количество ошибок при репликации [11].
4. Кодирующие CDS содержат гораздо больший процент GC по сравнению с некодирующими. Последние зачастую служат местом нейтральных мутаций, поэтому количество GC может сильно уменьшаться [12].
Благодарности
Благодарю своих бабушку и маму за то, что они готовили кушать, пока я изучал архею выше).
1. Dimitry Y. Sorokin, Ilya V.Kublanov, Mikhail M. Yakimov, W.Irene C. Rijpstra, Jaap S. Sinninghe Damste (2016), Halanaeroarchaeum sulfurireducens gen. nov., sp. nov., the first obligately anaerobic sulfur-respiring haloarchaeon, isolated from a hypersaline lake. Microbiology society
2. Axel Tiessen, Paulino Pérez-Rodríguez, Luis José Delaye-Arredondo (2012), Mathematical modeling and comparison of protein size distribution in different plant, animal, fungal and microbial species reveals a negative correlation between protein size and protein number, thus providing insight into the evolution of proteomes. BMC
3. Nobuhiko Tokuriki, Dan S Tawfik (2009), Stability effects of mutations and protein evolvability. ScienceDirect
4. Mark B. Gerstein, Can Bruce , Joel S. Rozowsky, Deyou Zheng, Jiang Du, Jan O. Korbel , Olof Emanuelsson, Zhengdong D. Zhang, Sherman Weissman, and Michael Snyder (2007), What is a gene, post-ENCODE? History and updated definition. Cold Spring Harbor Laboratory Press Bookstore (CSH)
5.ZhaoLei Zhang, Mark Gerstein (2004), Large-scale analysis of pseudogenes in the human genome. ScienceDirect
6.Cecilia Guerrier-Takada, Katheleen Gardiner, Terry Marsh, Norman Pace, Sidney Altman (1983), The RNA moiety of ribonuclease P is the catalytic subunit of the enzyme. 50 Cell
7. Christian Zwieb, Jerry Eichler (2001), Getting on target: The archaeal signal recognition particle. PMC
8. Christopher M. Thomas, Kaare M. Nielsen (2005), Mechanisms of, and Barriers to, Horizontal Gene Transfer between Bacteria. Nature
9. S Sáenz-Lahoya, N Bitarte, B García, S Burgui, M Vergara-Irigaray, J Valle, C Solano, A Toledo-Arana, I Lasa (2019), Noncontiguous operon is a genetic organization for coordinating bacterial gene expression. PMC
10. Alexander E Vinogradov (2003), DNA helix: the importance of being GC-rich. Oxford Academic
11. T Ikemura (1985), Codon usage and tRNA content in unicellular and multicellular organisms. Oxford Academic
12. N Sueoka (1993), Directional mutation pressure, mutator mutations, and dynamics of molecular evolution. Springer Nature